
|
|||
|---|---|---|---|
|
|
La potenciación post-activación muscular mediante estimulación eléctrica: una revisión |
|
|
|
Departamento de Educación Física y Deportiva. Universidad de Granada (España) |
Bernardo Requena Sánchez, Mikel Zabala Díaz, Cristóbal Sánchez Muñoz, Ignacio Chirosa Ríos y Jesús Viciana Ramírez mikelz@ugr.es |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 8 - N° 57 - Febrero de 2003 |
|
|
1 / 2
1. Introducción
Es comúnmente asumido que la actividad previa puede influir en el funcionamiento posterior del músculo esquelético (1). En este sentido, la activación muscular precedente a la contracción puede tanto disminuir la capacidad de rendimiento neuromuscular (produciendo fatiga y entendiendo ésta como un decrecimiento en la producción de fuerza -F- (5) como aumentarla. A este incremento en la F desarrollada como resultado de una activación previa se le denomina potenciación (1). La potenciación se desencadena a través de diferentes estrategias de activación (7): de forma endógena, inducida generalmente a través de pulsos de alta frecuencia inicial, fenómeno acontecido al inicio de movimientos balísticos (1); y de forma exógena o provocada. A esta forma exógena, se la denomina potenciación post-activación (PPA) (9).
Así, la presente revisión se centra sobre la PPA, y especialmente sobre la PPA inducida mediante estimulación eléctrica percutánea (EEP) (9, 10, 16, 17, 18). La totalidad de estudios que utilizan EEP como medio generador de PPA, obtienen mejoras significativas en diferentes gestos localizados. Fundamentalmente, estos gestos han sido monoarticulares y medidos mediante máquinas dinamométricas. El hecho de que la práctica totalidad de los estudios de rigor se hayan centrado en gestos localizados y, además, se hayan observado diferentes protocolos de potenciación, lleva a la necesidad de conjugar y aunar los aspectos clave de la PPA para tratar así de aplicar este fenómeno a gestos más complejos. Con esta revisión bibliográfica, se pretende ofrecer un punto de partida para posteriores estudios experimentales en gestos deportivos concretos.
2. Cuantificación de la PPAEl presente apartado se centra en los trabajos que han provocado y analizado la PPA en distintos grupos musculares del cuerpo humano (Tabla 1). En su totalidad, cuantifican la magnitud de la PPA comparando el comportamiento mecánico del grupo muscular objeto de estudio en los estados “potenciado” (o bajo el efecto de la PPA) y “no potenciado”.
2.1. Fases del proceso de inducción de la PPA.
La bibliografía consultada muestra diversos métodos para inducir la PPA (Tabla 1). En todos se pueden distinguir tres fases:
1ª fase. Evaluación del comportamiento mecánico del grupo muscular seleccionado en estado “no potenciado”.
2ª fase. Aplicación del estímulo desencadenante de la PPA. La PPA se evoca exógenamente mediante contracciones voluntarias de carácter máximo (2, 8, 11, 20, 27) o mediante estimulación eléctrica de carácter tetánico (9, 10, 16, 17, 18). En determinados trabajos en los que se emplean contracciones voluntarias, se comprueba que sólo es necesario un esfuerzo voluntario breve pero intenso para inducir PPA (3, 20, 27). O'Leary et al. (1997) señalan que una limitación del uso de contracciones voluntarias para provocar la PPA es la incapacidad de algunos sujetos de activar completamente sus músculos. Esto conlleva que no se alcance una potenciación máxima y se cause una mayor variación intersujeto (16, 17). Estas limitaciones no están presentes en los estudios en los que se emplean técnicas de EEP a un nivel máximo y constante (17), ya que, además de producir una activación completa del grupo muscular deseado, la EEP permite medir la capacidad de producción de F de los músculos independientemente de la voluntad y especialmente, del grado de motivación de los sujetos (20). Otro aspecto determinante en esta fase es la duración del estímulo inductor (Tabla 1); así, la mayor PPA inducida mediante una CMV, se alcanza mediante acciones isométricas de @ 10s de duración (27). En cambio, en el empleo de EEP tetánica, la mayor PPA se logra mediante corrientes de alta frecuencia mantenidas en un rango de 5 a 10s (9, 10, 17).
3ª Fase. Evaluación del comportamiento mecánico del grupo muscular en estado “potenciado”. En esta fase se realizan mediciones puntuales sucedidas escalonadamente en el tiempo, según el protocolo empleado. Con ello, se pretende describir cómo evolucionan los parámetros descriptores de la PPA en el tiempo. La magnitud de la PPA (determinada por los parámetros descriptores) decrece hasta los valores basales (propios del estado “no potenciado”) en un periodo de tiempo que oscila entre 5 y 20 min., con un tiempo medio de 10 min. (11, 17, 27) (Figura1).
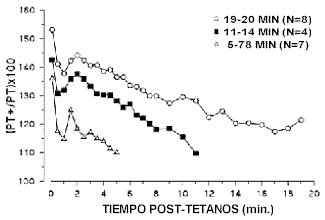
Figura. 1. Evolución de la magnitud de la PPA tras una contracción tetánica inducida a 100Hz, en tres grupos de sujetos (O'Leary et al., 1997). El valor de la PPA se expresa como porcentaje del valor de la misma en estado “no potenciado” (se utiliza el parámetro descriptor FIM que se tratará en los apartados siguientes). (Modificado de O’Leary et al. 1997).2.2. Grupos y acciones musculares implicados.
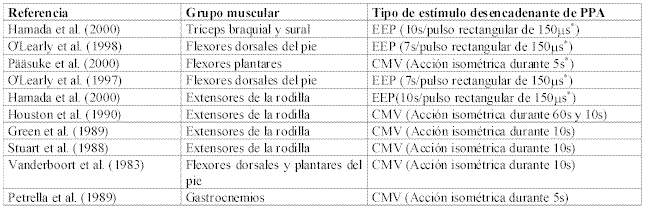
La Tabla 1 muestra la variedad de grupos musculares humanos en los que se ha medido la PPA y se completaría con los trabajos citados por O’Leary et al. (1997) en músculos de la cara y la mano. La selección de dichos grupos musculares responde al hecho de que cuando estos se activan (p.e. tríceps sural) desencadenan el movimiento de un segmento corporal (p.e. pie) cuyo comportamiento mecánico es cuantificable. La medición de la PPA en estos trabajos se ha llevado a cabo en base a acciones isométricas, de ahí que en numerosos trabajos se cuantifique la magnitud de la PPA con dinamómetros isométricos de fabricación propia (9, 10, 16, 17). En todos se emplea un transductor de F (células de carga) y soporte informático para la recogida y tratamiento de los datos. No se han encontrado trabajos referentes a la evaluación del efecto de la PPA sobre acciones de tipo concéntrico en humanos, aunque sí sobre pequeños mamíferos (1, 6).
Tabla 1. Principales trabajos en los que se induce la PPA, grupos musculares implicados y tipo de estímulo desencadenante seleccionado.
2.3. Parámetros descriptores de la magnitud de la PPA.
Teniendo en cuenta que se miden acciones isométricas, se deduce que el parámetro principalmente analizado para cuantificar la magnitud de la PPA, es la F isométrica máxima (FIM). Así mismo, en los trabajos que emplean la EEP como estímulo desencadenante de la PPA, se utilizan dos nuevos parámetros descriptores (9, 10, 16, 17, 18). Esto se debe al empleo de series de estímulos individuales de máxima intensidad (previamente fijada para cada individuo), que provocan el reclutamiento momentáneo de la totalidad de unidades motrices del grupo muscular en cuestión (máxima contracción tetánica) (17). Por consiguiente, se genera una acción isométrica máxima y aislada por cada serie de estímulos. Los dos nuevos parámetros antes mencionados se ubican en la curva F-tiempo de cada una de esas acciones isométricas (Figura 1). Algunos trabajos que emplean la MCV isométrica como estímulo desencadenante también aplican estos parámetros (2, 8, 20). El primer parámetro es el intervalo de tiempo transcurrido desde que comienza a generarse F hasta que la F isométrica máxima se alcanza (TFIM), y el segundo es el intervalo de tiempo que transcurre desde que se alcanza la FIM hasta que, por la relajación muscular que conlleva el cese de la activación, se llega a la mitad del valor de la FIM. A este intervalo se le denomina tiempo de relajación medio (TR1/2).
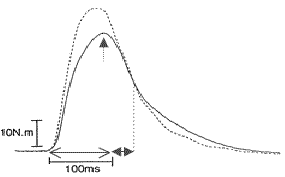
Fig.2. Ubicación de los tres parámetros descriptores de la magnitud de la PPA sobre la curva F-tiempo de una acción isométrica generada mediante EEP:FIM,
TFIM,
TR1/2. Figura modificada de Hamada et al. (2000).
2.4. Principales resultados.
La magnitud de la PPA se cuantifica en base a la comparación de los parámetros descriptores (FIM, TFIM, TR1/2), tanto en el estado "no potenciado" como "potenciado" del músculo objeto de estudio. En todos los trabajos recogidos, la tendencia de los parámetros descriptores es similar. En éstos, la PPA provoca un incremento estadísticamente significativo de la FIM y una disminución (generalmente significativa) en el TFIM y el TR1/2 (2, 8, 9, 10, 16, 17, 20). Hamada et al. (2000) muestran un incremento en la FIM de un 70´6% respecto a la FIM observada en estado "no potenciado" del cuadriceps de sujetos sedentarios. Por otro lado, O’Leary (1997) indica una disminución de un 14% y un 23% para el TFIM y el TR1/2 respectivamente, en referencia al TFIM y TR1/2 obtenido en estado "no potenciado" sobre los músculos flexores dorsales del pie. Estos ejemplos corresponden al valor de la PPA en los 5s posteriores de la 2ª fase del proceso de inducción de la PPA o fase del estímulo desencadenante.
Además, Hamada et al. (2000 a, b) muestran una correlación significativa y negativa entre la magnitud de la PPA, medida sobre el cuadriceps de 20 sujetos, y el valor del TFIM y TR1/2 en estado “no potenciado” (Figuras 3 y 4). Esto es, un TFIM y TR1/2 más corto (en estado “no potenciado”) fue asociado con un mayor PPA (9, 10, 17).
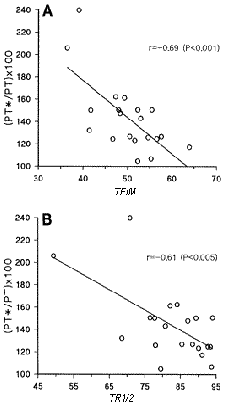
Figuras 3 y 4. Correlación entre la inmediata PPA (5s.) -medida mediante la FIM y
expresada como porcentaje de la FIM en estado “no potenciado"- y
el TFMI y TR1/2 en estado “potenciado”. (Tomado de O´Leary et al. 1997).
3. Mecanismos implicados en la PPADos teorías han intentado explicar la PPA en el músculo esquelético:
3.1. La fosforilación de la cadena ligera de miosina (CLM).
La incorporación de fosfato por la CLM ocurre cuando el Ca²+ liberado por el retículo sarcoplasmático forma el complejo Ca4+ -calmodulina activando una enzima específica, la kinasa de la cadena ligera de miosina (KCLM) (7, 13). Por consiguiente, el grado de participación de la KCLM depende de la frecuencia y duración de la activación muscular (7). De forma sucesiva, la magnitud de la fosforilación de la CLM dependerá tanto de la activación de la KCLM, como de la fosfatasa de la CLM (enzima desfosforiladora de la CLM perteneciente a la clase 1 de las proteínas fosfatasas (4, 23)). Manning y Stull (1982) fueron los primeros autores en establecer que la PPA generada por un músculo rápido de rata se correlaciona con el grado de fosforilación de 18.5KDa de MLC. Estos autores demostraron cómo tras una estimulación tetánica corta (1-2s) y de frecuencia elevada (200Hz), la fosforilación de la CLM incrementó desde unos niveles basales de 0.1-0.2 molP/mol CLM a 0.7 molP/molCLM a los 2s. Este nivel no se mantuvo constante y la fosforilación decreció lentamente hasta alcanzar el valor pretetánico o basal en 3-5 min., comprobándose una evolución paralela de la PPA. Así mismo, indicaron que la PPA es un proceso lento, pues alcanza su valor máximo a los 10-20s tras la tetania muscular. En esta línea, destaca el estudio de Sweeney y Stull (1990) quienes demostraron, sobre músculo esquelético de conejo, que la fosforilación de la cadena ligera de miosina provoca un aumento de la FIM y del ritmo de producción de la misma (relación F-tiempo).
En el músculo esquelético humano se ha constatado la fosforilación de la CLM en respuesta a contracciones máximas de corta duración (4, 11, 22). Aunque sobre el músculo vivo e intacto, los intentos por correlacionar la fosforilación de la CLM con los cambios que la PPA desencadena en la relación F-velocidad han generado resultados contradictorios (4). Sin embargo, los trabajos más recientes a este respecto han determinado una correlación significativa entre la elevada incorporación de fosfato por la CLM y la mejora en los valores obtenidos en la relación F-velocidad, como consecuencia de la potenciación (6). Así mismo, los estudios con músculos atrofiados aportan un nuevo argumento en favor de esta teoría (la fosforilación de la CLM como primer mecanismo responsable de la potenciación). Se conoce que la magnitud de la potenciación disminuye significativamente en músculos atrofiados (25, 26). Además, Tubman et al. (1997) comprobaron que, por un lado en el músculo gastrocnemio atrofiado de rata las CLM permanecen intactas y, por otro éstas se fosforilan en menor proporción que las CLM del grupo control en condiciones postetánicas. La asociación de estos dos hallazgos en el músculo atrofiado (de una fosforilación disminuida y una pérdida de PPA), proporciona una evidencia indirecta de que la fosforilación de CLM es el mecanismo principal de la potenciación y, por tanto de la PPA (22, 23).
| Lecturas: Educación Física y Deportes · http://www.efdeportes.com · Año 8 · Nº 57 | sigue Ü |