
|
|
|---|---|
|
Concepto y taxonomía de la fatiga física (II)
|
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 7 - N° 38 - Julio de 2001 | |
2 / 2
La fatiga central, aunque algunos autores le restan importancia, podría ser determinante para aquellos deportes de fuerza máxima y coordinación que exijan una gran activación neuromuscular.
Puntos de origen de la fatiga centralDurante la actividad muscular, la fatiga puede alterar los mecanismos de la contracción muscular a los diferentes niveles en los que se ponen en funcionamiento un proceso tan complejo como el que comprende desde que se inicia, a nivel del sistema nervioso central se propaga hasta el músculo y se realiza la contracción, apareciendo la respuesta motora, visible o no.
El origen se produce en uno o varios niveles de las estructuras nerviosas que intervienen en la actividad física. Responde sobre la alteración de la motivación, o bien por la alteración de transmisión de órdenes provenientes del SNC o de la captación de axones motores. Habitualmente se acepta su existencia asociada tanto a esfuerzos prolongados de baja intensidad como a esfuerzos de alta intensidad, sostenidos o en forma de repetición.
Green (1987) propone cuatro posibles puntos de aparición de la fatiga central:
Nivel supraespinal;
Inhibición aferente desde husos neuromusculares y terminaciones nerviosas;
Depresión de la excitabilidad de la motoneurona;
Fallos en la sinapsis.
Palomero y col. (1990, op. cit. en García Manso, 1996, pág.:43), a este nivel, proponen un mecanismo relacionado con la alteración en los neurotransmisores de las sinapsis entre las microneuronas del circuito de Renshaw y las alfa-motoneuronas. Este circuito, normalmente inhibidor, por acción del ácido gamma amino butírico (GABA) y la glicocola o glicina, puede llegar a modificar su acción transformándola en excitadora por acción del aspartato, glutamato y la acción reguladora de la taurina.
Fatiga periférica.
Definición, causas y orígenesSiguiendo la clasificación propuesta por Green (1990, Op. cit en Master C.O.E., Módulo, 5.2, pág.: 36) la fatiga podría afectar a las estructuras por debajo de la placa motora que intervienen en la contracción muscular (Figura 5):
Fallo pre-sináptico (Fatiga Central)
Dificultad para desarrollar el potencial de acción (liberación de Acetilcolina)
Fallo en el Sarcolema, para la propagación del impulso nervioso.
Fallo en el acople de los tubos T y el Retículo Sarcoplásmico.
Fallo en la liberación del Calcio con la Troponina.
Fallo en la afinidad del Calcio con la Troponina.
Fallo en la producción de puentes Actina-Miosina.
Fallo en el proceso de relajación.
Fallo en el proceso de relajación.
Figura 5. Lugares de aparición de la Fatiga Periférica. 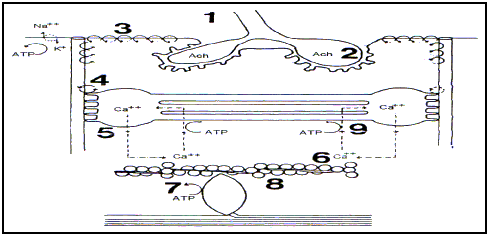
A nivel de placa motora (Puntos 1 y 2 de la Figura 5)Puede existir una disminución de la liberación del Neurotrasmisor (Acetilcolina). Así mismo puede existir un fallo en la propagación del potencial de acción por la acción de la Ach sobre la membrana postsináptica. Este punto de fatiga, en placa motora, se discute si es central o periférica. Su estudio se realiza mediante EMG, comprobando la amplitud y el área del potencial de acción.
Por lo tanto, a este nivel, se aprecia la alteración en la propagación del potencial de acción desde la vía nerviosa.
A nivel de sarcolema (véase punto 3, Figura 5)Ocurre lo mismo que a nivel de la membrana postsináptica de la placa motora. Puede existir una alteración en la propagación del potencial de acción debido a la salida de K+ intracelular y a la entrada de H2O. Esta reducción se trasmite a los TT y podría resultar en una disminución de la liberación de Ca++ desde el Retículo Sarcoplásmico.
El Sarcolema es una estructura activa que, mediante su permeabilidad, controla el metabolismo desde distintos puntos (Brooks, 1986):
Transporte de sustratos (d-glucosa, FFA, y proteínas).
Regulación por otras sustancias (insulina, receptores adrenérgicos, AMPC, IP3, Ca++).
Transporte de metabolitos tales como el lactato, etc..
En este último apartado es importante el transportador Lactato/H+. El Ácido láctico tiene un pH de 3.9, por lo que a pH normal está disociado en lactato y H+, siendo un transporte saturable (Km: 20 mM), mientras que el H+ tiende a sufrir un equilibrio iónico. Así se aprecia una alteración después de ejercicio supramáximo (Oyono-Enguelle y cols., 1992).
De este modo la alteración en el funcionamiento del sarcolema puede comprometer todas estas funciones.
Durante el ejercicio existe un flujo de Ca++, existiendo unos canales de entrada, dependientes del voltaje, estos canales están modulados por las catecolaminas (AMPc) y una bomba Na/Ca++. El influjo de Ca++ estimula la enzima Fosfofructoquinasa (PFK) y modula, posiblemente, la actividad de los canales de Potasio (K) que van a ser, en última instancia, determinante en la aparición de fatiga a este nivel.
En la Fatiga que afecta al sarcolema (Davies N.W., 1992, Op. Cit. en Marconnet et al., 1992, existe un fallo en el funcionamiento de la membrana, producido por las alteraciones de iones (K+i -intracelular-, K+e -extracelular-, Na+i y Cl-i), perdiendo el interior de la célula la polarización fisiológica. Fundamentalmente existe una salida de K+ (y entrada de Na+), ocasionando una disminución del K+i, dando una alteración del potencial de reposo de membrana. La recuperación, de la actividad normal de la membrana, es concomitante al proceso de recuperación del K+i, restableciéndose el potencial de membrana previamente al de los TT (en los que durante la fatiga existe un incremento del umbral de excitación). Se ha demostrado que la bomba Na/K está inhibida durante el proceso de Fatiga. El grado de pérdida de K+ es tres veces mas que la ganancia del Na+, debido a un aumento de la conductancia del K+ (por el aumento de Ca++i que aumenta la sensibilidad de los canales y por la deplección de ATP que activa dicha conductancia).
También es posible, que determinados canales para el K+ se abran por disminución de ATP, disminuyendo el potencial de reposo, así como la superficie y duración del potencial de acción.
Alteraciones del pH han sido reconocidas como mediador en la producción de la fatiga en ejercicios de alta intensidad, aunque existen situaciones de fatiga sin pH bajo.
Este se recupera mas despacio que el K+ o Na++ y está en relación con láctico/H+. El pH bajo también ocasiona una reducción (3 veces) en la conductancia del Ca++ en el RS.
En la fatiga que aparece en ejercicios de baja intensidad puede encontrarse explicación a las alteraciones de membrana motivadas por la activación de las fosfolipasas y/o producción de radicales libre durante el ejercicio. Al final puede inducir una peroxidación lipídica, en relación directa con las modificaciones de transporte de la membrana.
Por tanto, a nivel del sarcolema la fatiga estará condicionada por la pérdida de la actividad electrolítica de la membrana a causa de la salida de iones K+, como consecuencia de distintos mecanismo ocasionados por el ejercicio (pH, radicales libres, caída de ATP...).
A nivel del retículo sarcoplásmico (puntos 4 y 5, Figura 5)Durante la fatiga, se produce una disminución de Ca++ en cada onda de pulso, desde que ocurre la despolarización a partir de los TT y del sarcolema. Esta despolarización iónica puede sufrir el mismo mecanismo de fatiga (por alteración del K+i y del Na+e) que la membrana de la célula muscular.
El impulso eléctrico activa la liberación de Ca++ (punto 5, Figura 5) se lleva a cabo de la forma siguiente : el potencial de membrana de los TT ocasiona una transformación del PIP2 (fosfoinositol) a través de la Fosfolipasa C en DG (2x3 DiacilGlicerol) e IP3 (inositol 3 fosfato), siendo éste el encargado de activar los filamentos deslizantes del RS para la liberación del Ca++, siendo rápidamente degradado por las monoesterasas hacia Inositol. Estos productos son regenerados en PIP2 con un aporte de energía. Durante la fatiga hay una deplección de algunos de estos componentes (figura 6).
Se ha comprobado que la cafeína facilita la liberación de Ca++ desde el RS, por actuar directamente en los canales de liberación, aumentando la producción de fuerza.
La reducción de Ca++ es debido a la alteración en la liberación y/o a la rápida recaptación por el RS. A esto hay que añadir las proteínas citoplasmáticas que captan Ca++.
La reversibilidad de estos procesos se calcula tardan entre 24 y 48 horas.
Esta disminución del Calcio intracelular (Ca++i) va estar en relación con la tensión muscular desarrollada. Así, a mayor concentración de Ca++i mayor tensión desarrollada.
Por último, puede ser que, debido a la dependencia del ATP para la apertura de los canales del Retículo Sarcoplásmico, en situaciones de compromiso metabólico, pueda significar la energía para este proceso un factor limitante.
En resumen, a este nivel la fatiga altera la liberación del Ca++.
Unión del Ca++ con la troponina x c. (Punto 5, figura 5).La disminución en la capacidad de unión varía la interacción Actina-Miosina (AxM) ( puntos 7, 8 y 9,figura 8). En este sentido puede existir una disminución de la sensibilidad (de 1.7 veces) de estas proteínas al Ca++, como ocurre cuando aumenta los H+, ya que estos protones compiten por los receptores del Ca++. Siendo por tanto un mecanismo que contribuye a la fatiga.
En este punto la fatiga condiciona el nivel de afinidad del Ca++ con las proteínas contráctiles.
Interaccion actina-miosina (AxM), (puntos 7, 8 y 9, figura 5).Es necesario que se establezcan los puentes AxM, que son dependientes de energía.
Algunos metabolitos como el Pi producen una inhibición a éste nivel.
Es conocido que en presencia de Ca++, sin que existan H+, sin llegar a la presencia de acidosis del metabolismo anaeróbico, y la aparición de Pi, procedente de la ruptura de PCr y ATP, condiciona una menor tensión en la contracción muscular debido a que en su presencia no se produce acople entre la actina y la miosina.
La relajación (puntos 8 y 9, figura 5)
Ocurre cuando el Ca++i es recaptado al RS, por la disociación de los puentes de Actina-Miosina. Este proceso requiere energía para que la Bomba de Ca++ funcione. En estados de fatiga se observa acrecentamiento del Ca++i. Debido a que el aumento de Pi inhibe Ca ATPasa (Lewis y cols., 1985).
En este último punto, de interacción entre actina y miosina, será la presencia de sustancias (Pi) las que impidan la formación de puentes Actina-Miosina. Y será la alteración en la recaptación de Ca++ al retículo sarcoplásmico, los mecanismos de fatiga a este nivel.
Puede apreciarse que son muchos los posibles lugares de aparición de la fatiga, muchas veces zona de aparición dependerá del tipo de ejercicio-deporte que se realiza y su intensidad. Por lo que para un perfecto conocimiento de las situaciones de fatiga, el entrenador, técnico o docente debe conocer a la perfección su deporte y fisiología del mismo.
Orígenes de la fatiga periféricaTiene lugar en las estructuras que intervienen en la acción muscular y que se produce a niveles bajo de la placa motriz. Allen y col. (1992) o Vollestad y Sejersted (1988) sitúan los posibles puntos de producción de fatiga a este nivel en:
Pérdida en la velocidad de conducción del potencial de acción sobre la superficie de la fibra.
Modificación de la transmisión de la señal desde los tubos T al retículo sarcoplasmático.
Reducción en la liberación de calcio intracelular durante la actividad.
Decrecimiento en la sensibilidad al calcio de los miofilamentos (Ca++/Troponina).
Bajada de la tensión producida por los puentes de actina y miosina.
Figura 6. Principales puntos de aparición de la fatiga a nivel central. Adaptado y 
modificado de Edwars en García Manso y col. 1996, pág. 44.
Figura 7. Principales puntos de aparición de la fatiga a nivel periférico. Adaptado y 
modificado de Edwars en García Manso y col. 1996, pág. 44.
Bibliografía
Allen, D.G., Westerblad, H., Lee, J.A. Lännergren, J. (1992) : Role of excitation-contraccion cupling in muscle fatigue. En Sports Medicine. Nº 13. Vol 2. pp.: 116-126.
Asmussen, E. (1979): Muscle fatigue. En Medical Science and Sports Review, nº 11, pp.: 313-321.
Barbany, J.R. (1990): Fundamentos de fisiología del ejercicio y del entrenamiento. Barcelona. Barcanova.
Bigland-Ritchie, B.; Jones, D.A.; Hosking, G.P. y Edwards, R.H.T. (1979): Central and peripheral fatigue in sustained maximum voluntary contractions of human quadriceps muscle. En Clinical Science and Molecular Medicine, nº 54, pp.: 609-614.
Bigland-Ritchie, B.; Dawson, N.J.; Johanson, R.S. y Lippold, O.J.C. (1986): Reflex origin for the slowing of motoneurone firin rates in fatigue of human voluntary contractions. En Journal of Physiology, nº 379, pp.: 1380-1392.
Brooks, G.A. (1986): Lactate production during exercise: oxidizable substrate versus fatigue agent. En Macleod y col. Londres. Exercise. Benefits, limits and adaptations. pp.: 145-158.
Davies, N.W. (1992): ATP-Dependent K+ channels and other K+ channels of muscle:How exercise may modulate their activity. En Marconnet, P.; Komi, P.V.; Saltin, B y Sejersted, O.M. (1992): Muscle fatigue mechanism in exercise and training. En Medicine Sport Science vol. 34. 38-45.
Edwards, R.H.T. (1981): Human muscle function and fatigue. Human muscle fatigue: Physiologycal mechanism. Londres. Ed. Whelan.
Fernández, J.M. y Diego, A.M. (1993): Hormonas y ejercicio. En González Gallego, J. (1992): Fisiología de la actividad física. Interamericana-MacGraw-Hill. pp.: 95-128.
García Manso J.M., Navarro Valdivieso, M. y Ruiz Caballero, J.A., (1996): Bases teóricas del entrenamiento deportivo. Principios y aplicaciones. Madrid. Gymnos.
Garland, S.J., garner, S.H. y Mccomas, A.J. (1988): Relationship between numbers and frecuencies of stimuli in human muscle fatigue. En Journal Aplied Physiology, nº 65, pp.: 189-93.
Green, H.J. (1987): Neuromuscular aspects of fatigue. En Canadien Journal Sport Science, nº 12 (Supl.1) 000S-000S.
Hortobágyi, T.; Houmard, J.A.; Stevenson, J.R.; Fraser, D.D.; Johns, R.A. e Israel, R.G. (1993): The effects of detraining on power athletes. En Medicine Sports Exerxise, nº 25, vol. 8, pp.: 929-935.
Ikai, M. y Steinhaus, A.H. (1961): Some factors modifying the expresion of human strength. En Journal of aplied Physiology, nº 28, pp.: 157-163.
Koutedakis, Y.; Frichknecht, R.; Vrbová, G. Sharp, N.C.C: y Bdgett, R. (1995): Maximal voluntary quadriceps strength patterns in olimpic overtrained athletes. En Medicine Science Sport Exercise, vol. 27, nº 4, pp.: 566-572.
Legido, J.C. (1986): Fatiga y entrenamiento. III Jornadas Nacionales de Medicina en Atletismo. Pamplona. ANAMEDE, pp.: 109-120.
Lehmann, M.; Dickhuth, H.H:; Gendrisch y cols. (1995): Training-overtraining. A prospective study with experienced middle and long distance runners. En International Journal Sport Medicine, nº 12, pp.: 444-452.
Lewis, S.F.; Haller, R.G.; Cook, J.D. y Nunally, R.L. (1985): Muscle fatigue in McArdle's disease studied by 32-P NMR: Effect of glucose infusion. En Journal Applied Physiology, nº 59, pp.: 1991-1994.
Lewis, S.F.; Haller, R.G.; Cook, J.D. y Nunally, R.L. (1985): Muscle fatigue in McArdle's disease studied by 32-P NMR: Effect of glucose infusion. En Journal Applied Physiology, nº 59, pp.: 1991-1994.
Merton, P.A. (1954): Speculations on the servo-control of movement. En Malcolm, J.L. y Gray, J.A.B. (eds) CIBA Foundation Symposium. The Spinal Cord, pp.: 247-260. London. Churchill Livingstone.
Odagiri, Y.; Shimomitsu, T.; Iwane, H. y Katsumura, T. (1996): Relationships between exhaustive mood state and changes in stress hormones following and ultraendurance race. En International Journal Sport Medicine, vol. 17, nº 5, pp.: 325-331.
Oyono-Enguelle, S.; Freun, H.; Lonsdorfer, J. y Pape, A. (1992): Impaired lactate exchange and removal abilities after supramaximal exercise in humans. En Marconnet, P.; Komi, P.V.; Saltin, B y Sejersted, O.M. (1992): Muscle fatigue mechanism in exercise and training. En Medical Sport Science vol. 34.
Polkey, M.I.; Kyroussis, D.; Hamnegard, C.H.; Mills, G.H. Green, M y Moxhan, J. (1996): Quadriceps strength and fatigue assessed by magnetic stimulation of the femoral nerve in man. En Muscle and nerve, nº 19, pp.: 549-555.
Sahlin, K. y Seger, J.Y. (1995): Effects of prolonged exercise on the contractile properties of human quadriceps muscle. En European Journal Applied Physiology, nº 71, pp.: 180-186
Vollestad, N. y Sejersted, O.M. (1988): Muscle glycogen depletion patterns in Type I and subgroups of Type II fibres during prolonged severe exercise in man. En Acta Physiology Scandinavica, nº 122, pp.: 433-441.
Wood, J.; Furbush, F. y Bigland-Ritchie, B. (1987): Evidence for a fatigue-induced reflex inhibition of motoneurone firing rates. En journal of Neurophysiology, nº 58, pp.: 125-137.
| Inicio |
Concepto y taxonomía de la fatiga física (1ª parte)
Entrenamiento Deportivo
| |
|
|---|---|
|
revista
digital · Año 7 · N° 38 | Buenos Aires, Julio de 2001 |
|