
|
|||
|---|---|---|---|
|
|
Spirulina como fonte protéica na recuperação de ratos desnutridos: efeitos sobre o músculo esquelético |
|
|
|
Universidade Estadual Paulista, UNESP Campus Rio Claro - SP (Brasil) |
Cláudia Regina Alves Fabrício Azevedo Voltarelli Maria Alice Rostom de Mello faunesp8@yahoo.com.br |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 10 - N° 86 - Julio de 2005 |
|
|
1 / 1
Introdução
Os efeitos da fome sobre populações têm sido observados desde épocas remotas e descritos através dos tempos. Contudo, pesquisas realizadas utilizando métodos científicos para avaliação da desnutrição, sua prevalência, natureza e abrangência de seus efeitos e integração com outros fenômenos, só tiveram início em épocas recentes.
Em 1928, a antiga liga das nações, sob o patrocínio de sua organização de higiene, procedeu a inquéritos em diferentes países. Foram, então, divulgados relatórios sobre as condições alimentares no mundo, demonstrando que mais de 2/3 da humanidade viviam em permanente estado de fome (CASTRO, 1965).
Mais recentemente, baseada em pesquisas nutricionais realizadas em todos os continentes, a Food and Agriculture Organization of the United Nations (2001) relatou que, embora em termos percentuais, tenha havido decréscimo na incidência de desnutrição entre 1990 e 1999, em números absolutos 623,7 milhões de pessoas nesses continentes ainda são acometidas por essa carência nutricional.
No Brasil, em 1996, foram efetuados inquéritos sobre o estado nutricional da população de menores de 5 anos, em áreas urbanas e rurais em diferentes regiões (MONTEIRO, 2003). Em áreas urbanas da região Centro-Sul (inclui as regiões Centro-Oeste, Sudeste e Sul) por exemplo, a prevalência da desnutrição situou-se próximo a 4,6%, enquanto que em áreas urbanas das regiões Norte e Nordeste chegou aos 16,6 e 13,0%, respectivamente.
Há várias décadas a desnutrição protéica vem preocupando autoridades ligadas aos setores de saúde pública, principalmente a Organização Mundial de Saúde (OMS), cujos técnicos têm debatido a descoberta e o emprego de novas fontes protéicas bem como as normas a serem seguidas para seu uso na alimentação humana (MOURA, 1981).
Além da agricultura e da pecuária convencionais, outras fontes protéicas têm sido examinadas. Nesse sentido, microorganismos têm recebido atenção especial como fonte alternativa de proteína na dieta. O termo biomassa ou single cell protein (scp) foi proposto por Tannenbaum (1975) como um termo genérico para indicar fontes protéicas brutas ou refinadas, originárias de microorganismos.
Existem, pelo menos, duas razões para se pensar a respeito do papel da biomassa na alimentação humana: a velocidade de reprodução dos microorganismos e o grau de controle que pode ser exercido sobre suas condições de crescimento (MOURA, 1981). Entre vários organismos que vêm sendo estudados, a alga verde-azulada spirulina é considerada um microorganismo promissor para essa finalidade (TANNENBAUM, 1975).
A desnutrição protéica implica em comprometimento funcional de diversos sistemas e órgãos de seus portadores, entre eles a musculatura esquelética (WATERLOW & ALLAYNE, 1974; TORUN & CHEW, 1994). Algumas alterações são totalmente revertidas com a recuperação nutricional, enquanto outras, não.
Assim sendo, no presente estudo, visou-se avaliar os efeitos da spirulina como fonte protéica na recuperação de ratos submetidos à desnutrição protéica.
ObjetivoO presente estudo foi delineado para avaliar os efeitos da spirulina como fonte protéica durante a recuperação nutricional de ratos submetidos à desnutrição protéica sobre o crescimento e aspectos metabólicos do músculo esquelético (músculo sóleo). Para inferir sobre o crescimento foram avaliados os teores de proteína e DNA musculares. Como índices metabólicos, por ser a glicose importante substrato energético para a contração muscular, foram avaliadas captação e oxidação de glicose, síntese de glicogênio e produção de lactato.
Material e métodosAnimais
Foram utilizados ratos da linhagem Wistar, jovens (28 dias de idade, no início do experimento), provenientes do Biotério Central da Universidade Estadual Paulista (UNESP) - Botucatu/SP. Os animais permaneceram em gaiolas coletivas (5 ratos por gaiola) e foram alimentados com dietas semi-purificadas e água ad libitum bem como mantidos sob ciclo periódico claro e escuro de 12 horas à temperatura média de 25±2°C.
DietasForam empregadas dietas semi-purificadas, contendo 17% de proteína na forma de caseína, 6% de proteína na forma de caseína ou 17% de proteína na forma de spirulina, conforme descrito no quadro 1.
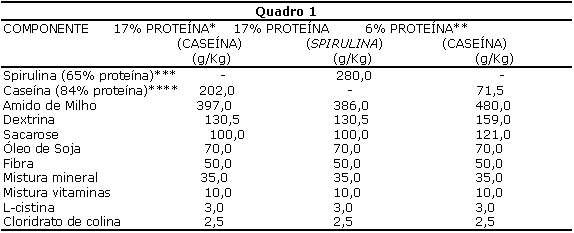
* Dieta para fase de crescimento, gravidez e lactação de roedores - AIN- 93G (REEVES et al., 1993).
** Dieta para indução de quadro de desnutrição (LATORRACA et al., 1998).
*** Valores corrigidos em função do conteúdo de proteína na spirulina.
**** Valores corrigidos em função do conteúdo de proteína na caseína.
Delineamento e grupos experimentaisO presente estudo foi delineado em duas etapas. Na primeira (fase de desnutrição), os ratos foram separados em dois grupos, de acordo com a dieta recebida dos 28 aos 60 dias de idade:
Grupo Controle - alimentados com dieta contendo 17% de proteína na forma de caseína
Grupo Desnutrido - alimentados com dieta contendo 6% de proteína na forma de caseína
Finalizado o período de desnutrição protéica, foi iniciada a 2 etapa (fase de recuperação nutricional), onde os ratos foram separados em 3 grupos, de acordo com a dieta recebida do 60 ao 90 dia de idade.
Grupo Controle - alimentados com dieta contendo 17% de proteína dos 28 aos 60 dias de idade e mantidos na mesma dieta dos 61 aos 90 dias.
Grupo Recuperado Caseína- alimentados com a dieta contendo 6% de proteína na forma de caseína dos 28 aos 60 dias e com a dieta contendo 17% de proteína na forma de caseína dos 61 aos 90 dias.
Grupo Recuperado Spirulina - alimentados com a dieta contendo 6% de proteína na forma de caseína dos 28 aos 60 dias e com a dieta com 17% de proteína na forma de spirulina dos 61 aos 90 dias.
ProcedimentosDurante todo o experimento, os animais foram pesados e medidos (comprimento focinho-ânus) semanalmente e tiveram a quantidade de alimento ingerida registrada diariamente. Ao final de cada etapa, os animais foram sacrificados por decapitação e foram coletadas: amostras de sangue para a determinação dos teores de proteína totais, albumina, ácidos graxos livres e glicose; fígado para a dosagem de lipídios e glicogênio e músculo sóleo esquerdo para a determinação dos teores de proteína e DNA. Foram ainda retiradas fatias do músculo sóleo direito, visando avaliar captação e oxidação de glicose, síntese de glicogênio e produção de lactato.
Metabolismo da glicose no músculo sóleoComo indicadores da funcionalidade do músculo esquelético foram avaliadas captação e oxidação de glicose, síntese de glicogênio e produção de lactato por fatias do músculo sóleo (25-35 mg) que foram incubadas por 60 minutos em tampão Krebs-Ringer bicarbonato suplementado com 5,5 mm de glicose, contendo [3H] 2-dioxiglicose (0,5mCi\ml) e [14C] -U- glicose (0,25mCi\ml) e com 100unm/ml da insulina. A captação e oxidação de glicose e a síntese de glicogênio foram determinadas pela medida de radioatividade do [3H] e do [14C] contidas, respectivamente, na fase alcalina e no precipitado obtido durante a extração do glicogênio. Para avaliar a produção de lactato, o substrato contido no meio de incubação foi separado por técnica de cromotografia de coluna (resina dowex-2) e a radioatividade do [14C] foi medida. Para detalhes referentes ao procedimento de incubação, consultar Prada et al. (2003).
CrescimentoVisando inferir quanto ao crescimento no músculo esquelético, foram determinados os teores de proteína e DNA (WINICK et al., 1972).
Dosagens Bioquímicas
Componentes sanguíneosOs teores séricos de proteínas totais, albumina, glicose e ácidos graxos livres foram determinados por métodos colorimétricos, conforme descrito por NOGUEIRA et al. (1990).
Proteína e DNA teciduaisOs teores de proteína no músculo foram determinados pelo método do folin-fenol (LOWRY et al., 1951) enquanto que os de DNA no mesmo tecido foram avaliados pelo método da difenilamina (GILES; MEYERS, 1965).
Glicogênio tecidualO glicogênio hepático foi extraído de acordo com procedimento descrito por SJORGREEN et al. (1938), e determinado pelo método do fenol (DUBOIS et al., 1956).
Análise EstatísticaOs resultados foram expressos como média ± desvio padrão e foram analisados por meio de teste t ou de análise de Kruskal-Wallis, onde apropriado. Em todos os casos o nível de significância foi de p<0,05.
ResultadosPrimeira etapa
Ao final da 1ª etapa, os ratos pertencentes ao grupo desnutrido apresentaram ganho de peso e comprimento corporais e concentrações de proteína total e albumina séricas (Tabela 1) e de glicogênio hepático significativamente menores quando comparados aos animais do grupo controle. Maiores concentrações de lipídios totais no fígado foram apresentados pelos mesmos animais (Tabela 1). As taxas de oxidação de glicose e síntese de glicogênio no músculo sóleo foram inferiores no grupo desnutrido se comparado ao grupo controle (Tabela 2). Não foram constatadas diferenças estatísticas entre os grupos nas concentrações musculares de proteína total e DNA musculares (Tabela 2).
Tabela 1. Ingestão alimentar (g/100g.dia) ao longo da 1ª etapa (desnutrição) do experimento; ganho de peso corporal (g) e incremento no comprimento corporal (cm); teores de proteínas totais (g/dl), albumina (g/dl), AGL ( Eq/l) e glicose(mg/dl) no soro dos animais; concentração de glicogênio (mg/10mg) e de lipídios totais (mg/100mg) no fígado dos animais ao final da 1ª etapa do experimento.
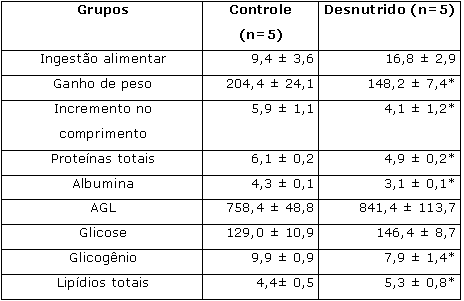
Resultados expressos como média ± desvio padrão;
* Diferença significativa (p < 0,05; teste t) em relação ao grupo controle.Tabela 2. Captação (mmol/g.h) e oxidação (mmol/g.h) de glicose; síntese de glicogênio (mmol/g.h); produção de lactato (mmol/g.h) e teores de proteína (mg/100mg de tecido) e DNA (mg/100mg de tecido) no músculo sóleo dos animais ao final da 1ª etapa (desnutrição) do experimento.
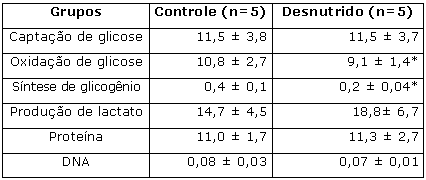
Resultados expressos como média ± desvio padrão;
* Diferença significativa (p< 0,05, teste t) em relação ao grupo controle.
Segunda etapaAo final da 2ª etapa, a ingestão alimentar apresentada pelos grupos recuperados caseína e spirulina foi significativamente maior em relação aos grupos controle (Tablea 3). Os ratos pertencentes aos grupos recuperado caseína e recuperado spirulina apresentaram ganho de peso e comprimento corporal superiores ao grupo controle (Tabela 3). Foi observada diferença significativa entre os grupos controle, recuperado caseína e recuperado spirulina quando analisado o teor de glicose no soro (Tabela 3), sendo os valores dos dois últimos superiores aos do primeiro. Para todas as outras análises realizadas no soro (Tabela 1) e nos tecidos (músculo sóleo e fígado - Tabelas 3 e 4) foram obtidos resultados semelhantes quando comparados os três grupos experimentais.
Tabela 3. Ingestão alimentar (g/100g.dia) ao longo da 2a etapa (recuperação) do experimento; ganho de peso corporal (g) e incremento no comprimento corporal (cm); teores de proteínas totais (g/dl), albumina (g/dl), glicose (g/dl) e AGL (µEq/L) no soro dos animais; concentração de glicogênio (mg/100mg) e de lipídios totais (mg/100mg) no fígado dos animais ao final da 2a etapa (recuperação) do experimento.
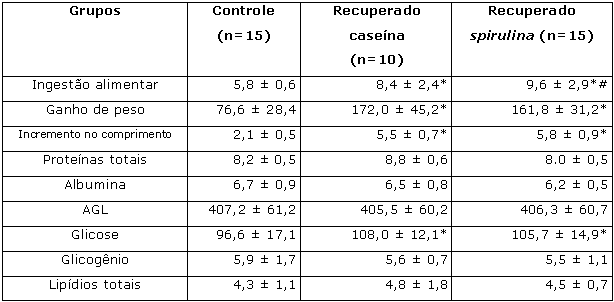
Resultados expressos como média ± desvio padrão.
* Diferença significativa (p< 0,05; Kruskal-Wallis) em relação ao grupo controle;
# Diferença significativa (p< 0,05; Kruskal-Wallis) em relação ao grupo recuperado caseína.Tabela 4. Captação (mmol/g.h) e oxidação (mmol/g.h) de glicose; síntese de glicogênio (mmol/g.h); produção de lactato (mmol/g.h) e teores de proteína (mg/100mg) e DNA (mg/100mg de tecido) no músculo sóleo dos animais ao final da 2a etapa (recuperação) do experimento.
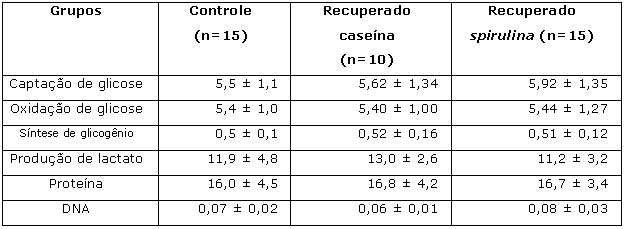
Resultados expressos como média ± desvio padrão.
DiscussãoA desnutrição protéica é considerada, ainda hoje, grave problema médico-social nos países em desenvolvimento.
Assim, é de interesse o desenvolvimento de procedimentos mais efetivos no seu tratamento. Isso inclui o emprego de novas fontes de proteína na alimentação, como por exemplo, a spirulina, uma alga verde-azulada de forma helicoidal, com comprimento de 0,2-0,5mm (BELAY et al., 1993).
O mérito especial dessa alga como fonte alimentícia é que a mesma contém 65 a 70% de proteína, com base no peso seco, o que é superior a qualquer outra fonte natural de alimento e contém os oito aminoácidos essenciais ao homem (NAKAYA et al., 1998). A alga cresce, normalmente, em águas de lagos naturalmente alcalino, localizados em zonas áridas. Embora a água alcalina desses lagos não possa ser utilizada para irrigação, pode ser aproveitada para o cultivo de spirulina e, uma vez que essa alga tem uma taxa de reprodução rápida, dividindo-se três vezes ao dia, uma área devotada totalmente ao crescimento de spirulina pode produzir 125 vezes mais proteína que área de mesma dimensão devotada ao cultivo de milho; 70 vezes mais do que se voltada à criação de peixes ou 600 vezes mais se voltada para criação de gado (FURST, 1978).
De acordo com avaliações prévias efetuadas com seres humanos, a spirulina parece ser boa fonte protéica para a alimentação, possui boa aceitação e não apresenta efeitos tóxicos aparentes (KAY, 1991). Em estudos conduzidos com animais de laboratório, especialmente ratos, constatou-se que, também para estes, a spirulina apresenta boa aceitação, adequada digestibilidade e ausência de efeitos tóxicos (SALAZAR et al., 1998).
Grinstead et al. (2000) observaram que uma dieta suplementada com 2,8 gramas de spirulina 3 vezes ao dia ao longo de 4 semanas resultou em significativa redução do peso corporal de porcos obesos. A spirulina demonstrou eficácia, também, em atenuar a hipertensão arterial de ratos (IWATA et al., 1990; HERNÁNDEZ et al., 2001). Além disso, outros trabalhos mostraram que a spirulina pode exercer efeitos benéficos no tratamento de algumas formas de câncer (COMBS et al., 1989; NAGASAWA et al., 1989) e de desnutrição (FICA et al., 1984).
Os resultados da 1ª etapa do estudo referentes ao ganho de peso e incremento de comprimento corporal, concentrações séricas de proteínas totais e albumina bem como teores hepáticos totais de glicogênio e lipídios, indicam que a alimentação dos animais com a dieta deficiente em proteína (6% caseína) foi eficaz em causar quadro de desnutrição semelhante ao Kwashiorkor infantil em seres humanos, exceto pela ausência de edema (GALDINO et al., 2001). A ausência de edema nos animais não surpreendeu, uma vez que outros pesquisadores também apresentaram dificuldade em produzir edema em animais com desnutrição protéica experimental (GALDINO, 1996).
A redução do ritmo de ganho de peso e crescimento linear tem sido relacionada às alterações funcionais e morfológicas, principalmente do pâncreas exócrino e do epitélio intestinal, que comprometem a digestão e a absorção dos nutrientes (GALDINO et al., 2001). Essas alterações observadas tanto no crescimento quanto na bioquímica sanguínea e tecidual dos ratos do presente estudo provavelmente foram ocasionadas pela deficiência protéica, uma vez que as dietas normo e hipoprotéicas eram isocalóricas e a quantidade de alimento ingerida pelos animais submetidos à dieta hipoprotéica foi maior do que daqueles submetidos à dieta normoprotéica durante o experimento.
No presente estudo, em sua primeira etapa, constatamos redução tanto na oxidação de glicose quanto na síntese de glicogênio pelo músculo esquelético dos ratos submetidos à restrição protéica, incubado na presença de insulina. Por outro lado, CRACE et al. (1995), CARNEIRO et al. (1995) e REIS et al. (1997), verificaram maior sensibilidade à ação da insulina na metabolização da glicose em animais tratados com dieta deficiente em proteína. CRACE et al. (1995) relataram maior captação de glicose em músculo sóleo isolado de ratos com restrição protéica em relação aos controles. Além disso, CRACE et al. (1990) verificaram aumento na síntese de glicogênio na musculatura de ratos com restrição protéica. Em outro estudo, NOLTE et al. (1994) não constataram diferença significativa entre animais controles e desnutridos quanto à síntese de glicogênio em fatias musculares incubadas na presença e na ausência de insulina. No entanto, esses autores verificaram redução na taxa de oxidação de glicose nas fatias musculares provenientes de animais desnutridos incubados na presença de insulina, como foi semelhantemente observado no presente estudo. As discrepâncias existentes entre os estudos provavelmente são devidas às diferenças nos protocolos experimentais utilizados.
Ao final da 2a etapa, os níveis séricos de proteínas totais, albumina e ácidos graxos livres, bem como as taxas de oxidação de glicose e de síntese de glicogênio no músculo sóleo incubado e concentrações hepáticas de glicogênio e de lipídios totais, foram restauradas pela realimentação com ambas as dietas, caseína e spirulina, uma vez que os ratos pertencentes aos grupos recuperado spirulina e recuperado caseína apresentaram valores semelhantes aos dos animais do grupo controle para essas variáveis metabólicas. Ganho de peso e incremento corporais foram maiores nos animais recuperados com ambas as fontes protéicas em relação ao grupo controle. Isso indica a eficácia das duas fontes protéicas na tentativa de recuperação desses parâmetros. Vale ainda ressaltar, que a ingestão alimentar ao final da 2a etapa mostrou-se significativamente maior no grupo recuperado spirulina quando comparado com os outros dois grupos do presente estudo. Tal fato evidencia a aceitação, por parte dos animais, da spirulina com fonte protéica na dieta administrada para a recuperação nutricional dos mesmos.
Os resultados encontrados ao final da 2a etapa, mostram-se coerentes com relatos anteriores da literatura, que apontam normalização de parâmetros bioquímicos sanguíneos e musculares bem como aceleração do crescimento somático após a realimentação de ratos previamente desnutridos com dietas balanceadas (GALDINO et al., 2001). Além disso, a dieta à base de proteína proveniente da spirulina mostrou-se tão eficaz quanto à dieta a base de proteína proveniente do leite (caseína) na recuperação nutricional dos animais.
Tais resultados corroboram observações prévias de que a spirulina, como única fonte de proteína, é adequada à promoção do crescimento somático e à manutenção da homeostase em ratos (CONTRERAS et al 1979, MARANES et al 1984, TRANQUILLE et al 1994). O fato de a spirulina ter-se mostrado tão eficiente quanto à caseína no processo de reabilitação da desnutrição protéica é de extrema importância, pois trata-se de fonte protéica de custos mais reduzidos do que a proteína de origem animal. Nesse sentido, é importante destacar que estudos adicionais em desenvolvimento em nosso laboratório indicam que a alimentação com spirulina promove maior estimulação na síntese protéica no músculo sóleo isolado do que a alimentação com caseína, em ratos eutróficos (VOLTARELLI et al., 2003). Estudos futuros são necessários para a verificação da aplicabilidade de dietas à base de proteína da alga spirulina na recuperação da desnutrição protéica.
ConclusãoEm conjunto, os resultados do presente estudo sugerem que:
A realimentação de ratos previamente submetidos à restrição protéica com dietas balanceadas, mostrou-se eficaz na reversão das alterações induzidas pela restrição;
Ambas as dietas empregadas para a recuperação nutricional (caseína e spirulina) mostraram-se igualmente eficientes;
Tal fato demonstra o potencial valor da proteína proveniente da alga na recuperação da desnutrição protéica;
Estudos futuros são necessários para verificação da aplicabilidade dessa proteína em dietas para recuperação nutricional.
Referências bibliográficas
BELAY, A; OTA, Y; MIYAKAWA, K; SHIMATSU, H. Current knowledge on potential health benefits of spiruline. J. Appl. Physic. 5: 235-241, 1993.
CARNEIRO, E.M; MELLO, M.A. R; GOBATTO, C.A; BOSCHERO, A. C. Low protein diet impairs glucose - induce insulin secretion from and 45Ca uptake by pancreatic rat islets. Nut. Biochem. 6: 314 - 8, 1995.
CASTRO, J. Geopolítica da fome. 7a ed., Rio de Janeiro, Editora Brasiliense. vol. 1, 54-55, 1965.
COMBS, W; SONIS, S.T; FITZGERALD, J; TRACY, C; WILSON, R. In vivo and in vitro effects of beta carotone and algae extracts in murine tumor models. Nutr. Cancer. 12: 371, 1989.
CASTRO, J. Geopolítica da fome, 7°ed, Rio de Janeiro, Brasiliense, 1965, V. 1, P. 54-5.
CONTRERAS, A; HERBERT, C.; GRUBBS, B. G., CAMERON, I. L. Blue-green alga spirulina as the sole dietary source of protein in sexually maturing rats. Nutr. Rep. Int 19: 749-763, 1979.
CRACE, C.J; SWENNE, I; KHON, P.G; STRAIN, J; MILNER, R. D.G. Protein energy malnutrition induces changes in insulin sensitivity. Diabetes Metabolism. 16: 484-491, 1990.
CRACE, C.J; SWENNE, I; KHON, P.G; STRAIN, J; MILNER, R. D.G. Protein energy malnutrition induces changes in insulin sensitivity. Diabetes Metabolism. 16: 484-491, 1995.
DUBOIS, B. et al. Colorimetric method for determination of sugar and related substances. Anal. Chem. 18:350-6, 1956.
FICA, V; OLTEANU, D; OPRESCU, S. Use of spiruline as an adjuvant nutrient factor in the therapy of the diseases accompanying a nutritional deficiency. Rev. Med. Int. Med. Int. 36: 225, 1984.
FOOD AND ORGANIZATION OF THE UNITED NATIONS. Assessment of the World Food Security Situation. Committee on World Food Security Twenty-Seventh Session, Rome, May 28, June 1, 2001.
FURST, P.T. Spiruline - a nutritious alga, once, a staple of Aztecdiet, could feed many of the words hungry people. Human Nature. 60: 459-470, 1978.
GALDINO, R. Atividade física e ingestão da dieta hipoprotéica durante a gestação em ratas: efeitos sobre a gestante e o desenvolvimento fetal. Rio Claro-SP. Dissertação (Mestrado em Ciências da Motricidade) - Instituto de Biociências, Universidade Estadual Paulista, 1996.
GALDINO, R; MELLO, M.A.R; ALMEIDA, R.L; ALMEIDA, C.C.S. Desnutrição protéico-calórica. In: DÂMASO, A. Nutrição e exercício na prevenção de doenças. v.1, MEDSI, 225-275, 2001.
GILES, K.W; MEYERS, A. An improved diphelinanine method for the estimation of dioxinucleic. Nature. 206: 93-115, 1965.
GRINSTEAD, G.S; TOKACH, M.D; DRITZ, S.S; GOODBAND, R.D; NELSSEN, J.L. Effects of spirulina plantesis on growth performance of weanling pigs. Animal Feed Sci. Techn. 83: 237-247, 2000.
HERNÁNDEZ, A.R; CASTILLO, J.L.B; OROPEZA, M.A.J; ZAGOYA, J.C.D. Spirulina maxima prevents fatty liver formation in CD-1 male and female mice with experimental diabetes. Life Sci. 69: 1029-1037, 2001.
IWATA, K; INAYAMA, T; KATO, T. Effects of spirulina plantesis on plasma lipoprotein lipase activity in fructose-induced hyperlipidemic rats. J. Nutr. Sci. Vitamiol. 36: 165-171, 1990.
KAY, R.A. Microalgae as food and supplement. Clin. Rev. Food. Sci. Nutr. 30(6): 555-573, 1991.
LATORRACA, M.Q; GOBATTO, C.A; CARNEIRO, E.M; GALDINO, R.S; SIBUYA, C.Y; MELLO, M.A.R. Descrição de dieta purificada para indução de quadro de desnutrição protéica em ratos. Rev. Bras. Med. Esporte. 4: 9-12, 1998.
LOWRY, O.H; ROSEBROUG, N.J; FARR, A.L; RANDALL, R.T. Protein measurement with the folinphenol reagent. J. Biol. Chem. 193: 265-275, 1951.
MARANES, M; BARZANTI, V; CARENINE, G; GENTILI, P. Nutritional studies on spirulina maxima. Acta vitaminol Enzimol 6:295-304, 1984.
MONTEIRO, C.A. A dimensão da pobreza, da desnutrição e da fome no Brasil: implicações para políticas públicas. Seminário Especial Fome e Pobreza, Rio de Janeiro, Setembro/2003.
MOURA, E.C.V. Fontes protéicas não convencionais: perspectivas de seu emprego na alimentação. Nóbrega, F.J. Desnutrição intra-uterina e pós-natal. São Paulo, Panamed, 35-56, 1981.
NAGASAWA, H; KONISHI, R; SENSUI, N; YAMAMOTO, K; BEN-AMOTZ, A. Inhibition by beta carotone rich algae Dunaliella of spontaneous mammary tumourigenesis in mice. Anticancer Res. 9: 71, 1989.
NAKAYA, N; HOMMA, Y; GOTO, Y. Colesterol lowering effect of spiruline. Nutr. Rep. Intern. 37: 1329-1337, 1998.
NOGUEIRA, D.M; STRUFALDI, B; HIRATA, M.H; ABDALLA, D.S.P; HIRATA, R.D.C. Métodos de bioquímica clínica, São Paulo: Pancas Editorial, 1990.
NOLTE, L.A; GALUSKA, D; MARTIN, I.K; ZIERATH, J.R; WALLBERG-HENRIKSSON, H. Elevated free fatty acid levels inhibit glucose phosphorylation in slow-twitch rat skeletal muscle. Acta Physiol. Scand. , 151: 51-9, 1994.
PRADA, F.J.A; MACEDO, D.V; MELLO, M.A.R. Evaluation of a protein deficient diet in rats through blood oxidative stress biomarkers. Molec. Patol. and Pharmacol. 113(114): 213-227, 2003.
REEVES, P.G; NIELSEN, F.H; FAHEY, G.C. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition AdHoe Writing Committee on the Reformulation of AIN-76A. J. Nutr. 123: 1938-1951, 1993.
REIS, M.A. B; CARNEIRO, E.M; MELLO, M.A.R; BOSCHERO, A. C; SAAD, M. J; VELLOSO, L.A. Glucose induced insulin secretion is impaired and insulin phosphorylation of the insulinreceptor and insulin receptor substrate - I are increased in protein - deficient rats. J. Nutr. , 127: 403-10, 1997.
SALAZAR, M; MARTÍNEZ, E; MADRIGAL, E; RUIZ, L.E; CHAMORRO, G.A. Subchronic toxicity study in mice fed spiruline maxima. J. Ethnopharm. 62: 235-241, 1998.
SJORGREEN, B; NORDEN, S. K; JOLD, T; HOLMGREN, H; WOLLERSTRON, J. Bertrag zur kenthis des le berrhythmik. Pflügers. Arch. Gesante Physiol. Menscen Tiere. 240-247, 1938.
TANNENBAUM, S.R. Single cell protein. v.2, MIT, Massachussets, 40-78, 1975.
TORUM, B; CHEW, F. Protein energy malnutrition. In. Shils M, Olson JA, Shike M (eds). Modern Nutrition in Health and Disease. 2: Lea & Febiger: Philadelphia; 950-976, 1994.
TRANQUILLE, N; EMEIS, J.J; CHAMBURE, R; TAMPONNET, C; Spirulina acceptability trials in rats. A study for the "melissa" Life-support system. Adv. Space res. 14(11): 167-170, 1994.
VOLTARELLI, F.A; MELLO, M.A.R; MARCONDES, M.C.C.G. Spirulina protein, body growth and skeletal muscle protein metabolism in young rats. Amino Acids. 35: 150, 2003.
WATERLOW, D.C; ALLEYNE, G.A.O. Má nutrição em crianças: evolução dos conhecimentos nos últimos 10 anos. Nestlé, São Paulo, 61-67, 1974.
WINICK, M; BASEL, J.A; ROSSO, P. Nutrition and cellular growth. In: Winick, M. Nutrition and development, John Willey, 49-97, 1972.
| |
|
|---|---|
|
revista
digital · Año 10 · N° 86 | Buenos Aires, Julio 2005 |
|