
|
|||
|---|---|---|---|
|
|
Efeitos dos treinamentos de endurance e intervalado em marcadores do metabolismo oxidativo, oxidantes, antioxidantes e lesão muscular Endurance and intermittent training induce different adaptations on blood and muscle oxidative, oxidant, antioxidant and injury biomarkers |
|
|
|
* Laboratório de fisiologia do exercício, Faculdade Social da Bahia (FSBA). ** Laboratório de bioquímica do exercício (LABEX), Departamento de Bioquímica, IB, Universidade Estadual de Campinas (UNICAMP). (Brasil) |
Dr. Claudio Cesar Zoppi* Dr. Armindo Antonio Alves** Dr. Leonardo dos Reis Silveira** Dra. Lucia Pereira da Silva** Dra. Denise Vaz de Macedo** czoppi@fsba.edu.br |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 11 - N° 99 - Agosto de 2006 |
|
|
1 / 1
Introdução
O treinamento esportivo se baseia no princípio da adaptação, onde as cargas de treinamento são responsáveis pela quebra da homeostase intracelular e assim, capaz de sinalizar as adaptações desejadas (BRUIN et al., 1994; KUIPERS, 1998). O estresse oxidativo, condição na qual a produção de espécies reativas de oxigênio (EROs) supera a capacidade antioxidante intracelular de eliminá-los, é uma das formas de estresse induzida pelo treinamento esportivo (SJODIN et al., 1990; JI, 1999). Nesse sentido, estudos mostram a produção de espécies radicalares, tais como radical ânion superóxido (-O2.), peróxido de hidrogênio (H2O2) e radical hidroxila (OH.) durante a contração muscular (DAVIES et al. 1982; POWERS & LENNON, 1999).
O aumento do fluxo de elétrons da cadeia respiratória mitochondrial parece estar diretamente envolvido nesse processo, bem como o aumento na concentração do substrato da enzima xantina oxidase (SJODIN et al., 1990; JI, 1999; JI & LEICHTWEISS, 1997; TIIDUS, 1998). Outros autores também mostram que a alta atividade dos neutrófilos, macrófagos e citoquinas presentes no processo inflamatório, responsável pela remoção e reparo de estruturas celulares danificadas após o exercício físico, também contribuem para a produção de EROs (CANNON et al., 1991; FIELDING et al., 1993; HESSEL et al.,2000; SEN, 1995).
Dados da literatura mostram que as EROs estão relacionadas a danos oxidativos a estruturas intracelulares tais como; lipídeos de membrana, proteínas e inclusive o DNA (SJODIN et al., 1990; DAVIES et al. 1982; HALLIWELL & GUTTERIDGE, 1998). Assim, especula-se que as EROs possam desencadear as lesões musculares (JI, 1999; TIIDUS, 1998; HESSEL, 2000). O estresse imposto às células musculares pode levar a dois processos distintos: adaptação ou "overtraining" (JACKSON, 1999; SMITH, 2000; SMOLKA et al., 2000). Adaptação e a conseqüente melhora no desempenho esportivo acontecem quando o estresse é seguido por um tempo de regeneração adequado, através do processo conhecido como supercompensação (BRUIN et al., 1994; VIRU, 1984; KUIPERS, 1998). O treinamento de alta intensidade parece ser a melhor forma de melhorar o rendimento esportivo (ALESSIO, 1993; CRISWELL et al., 1993). No entanto, estar bem adaptado significa ter as fibras musculares aptas a suportar o estresse imposto pelo exercício, isto é, sua intensidade, duração e freqüência, fatores que podem resultar em fadiga e ainda levar a lesões musculares (POWERS & LENNON, 1999). A imposição de cargas de treinamento que excedem a capacidade do atleta de se regenerar leva ao aparecimento prematuro dos processos de fadiga. Tal estratégia, quando adotada de forma crônica, pode levar à instalação do processo inicial do "overtraining", denominado pela literatura de "overreaching" (KUIPERS, 1998; GLEESON, 2000).
Desde os primeiros estudos que propuseram o exercício físico extenuante como indutor de danos oxidativos em vários tecidos (DAVIES et al., 1982; DILLARD et al., 1978), um considerável corpo de conhecimento vem-se acumulando também a respeito das adaptações induzidas pela atividade física nos sistemas de defesa antioxidante (POWERS & LENNON, 1999; SEN, 1995; LIU et al., 2000). No entanto, é preciso destacar que ainda se conhece muito pouco a respeito da taxa de regeneração, processos adaptativos e quantidade ideal de estímulos de alta intensidade que devem ser aplicados nos programas de treinamento. Recentemente, demonstramos que os treinamentos de endurance (longa duração, alta intensidade) e intervalado (curta duração, alta intensidade) induzem adaptações diferenciadas sobre as enzimas citrato sintase (CS), glutationa redutase (GR) e catalase (CAT) no músculo sóleo (SMOLKA et al., 2000). Neste estudo, mostramos que o treinamento de endurance (CT) foi mais eficiente em aumentar a atividade das enzimas antioxidantes, enquanto o treinamento intervalado (IT) se mostrou mais eficiente no aumento do metabolismo oxidativo, medido pela CS (SMOLKA et al., 2000). Tais dados sugerem fortemente que a taxa de estresse oxidativo pode ser modulado pela intensidade, duração e tipo de exercício (SEN, 1995; SMOLKA et al., 2000). Além disso, ALESSIO e colaboradores (2000) também demonstraram a existência de um limiar de esforço, a partir do qual o estresse oxidativo aumentaria dramaticamente. Segundo os autores, esse limiar parece estar relacionado com o protocolo de treinamento e as condições individuais do atleta.
Neste estudo, seguimos o perfil adaptativo de biomarcadores de estresse oxidativo, enzimas do sistema de defesa antioxidante e do metabolismo oxidativo no sangue e também no músculo sóleo de ratos, por oito semanas, de dois protocolos distintos de treinamento, CT e IT. O objetivo foi verificar se os biomarcadores escolhidos seriam capazes de detectar o grau de impacto produzido pelos protocolos de treinamento no músculo, antecipando, ou não, o sucesso do processo adaptativo.
Materiais e métodosAnimais: Os experimentos foram conduzidos utilizando ratos da linhagem Wistar com dois meses de idade. Os animais foram mantidos em ciclo invertido de luz 12h claro-escuro, a 22o C sendo oferecido durante todo o experimento, água e ração específica para ratos à vontade. Todos os procedimentos experimentais foram aprovados pelo comitê de ética para o uso de animais em pesquisa do instituto de biologia da UNICAMP.
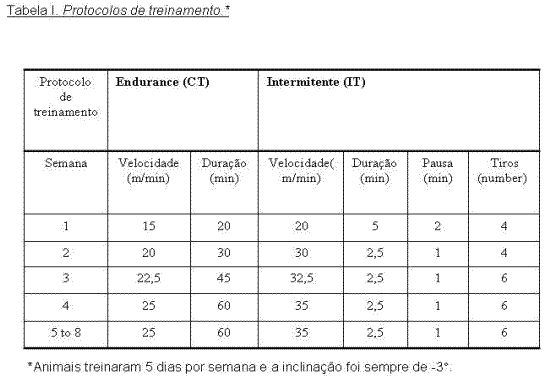
Protocolos de treinamento. Vinte e quatro ratos que se mostraram aptos a correr em esteira elétrica foram distribuídos ao acaso entre os grupos experimentais sedentários (SED), CT e IT, contendo cada grupo um n de 8 animais cada. As características dos protocolos de CT e IT estão mostrados na tabela I. Ambos os protocolos se apresentam divididos em duas fases com duração de um mês cada. A Fase I onde velocidade e duração aumentam gradativamente, e a Fase II onde os mesmos parâmetros permanecem constantes. Esta estratégia foi utilizada para simular um processo de treinamento, que contém um período de aquisição e outro de manutenção.
Quarenta e oito horas após o final da última sessão de treino, os animais foram sacrificados e as amostras de sangue e músculo dos animais dos grupos IT e CT coletadas. Os animais do grupo SED foram sacrificados e as amostras coletadas após as oito semanas do período de treinamento, a fim de evitar alterações induzidas pelo tempo nas variáveis estudadas.
Para dosagem de lactato, hemoglobina (Hb) e atividade da enzima creatina quinase (CK) (EC. 2.7.3.2) plasmática foram coletadas 150 µL de sangue da calda dos animais semanalmente, imediatamente após o término da última sessão de treinamento da semana. Esses três parâmetros foram dosados por espectrometria de refletância através de fitas reativas específicas, usando os aparelhos Accusport (FELL et al., 1998; BISHOP, 2001), no caso do lactato e Reflotron a hemoglobina e atividade da CK.
Preparação das amostras de músculo. Os Animais foram anestesiados com uma injeção intraperitoneal de hidrato de cloral 10% (p/v), com 0,3 mL por 100g de peso. Foram retirados os músculos sóleo de ambas as pernas, cuidadosamente dissecados e imediatamente congelados em nitrogênio líquido e mantidos a -70o C até que as análises fossem conduzidas. Os animais então foram sacrificados por decapitação. Trinta miligramas de amostra dos músculos foram picados em pequenos pedaços com tesoura e homogenizados em 2 mL de meio frio (8o C), contendo 440 mM sacarose, 50 mM MOPS, 0,01 mM PMSF e 100 mM EDTA, pH 7.2. Para a preparação do homogenato, foi utilizado o aparelho Polytron em sua maior velocidade por 30 segundos. Neste homogenato foram dosadas as atividades das enzimas CS (EC 4.1.37), GR (EC 1.6.4.2) e CAT (EC 1.11.1.6). Essas enzimas foram escolhidas por mostrarem modulação induzida pelo treinamento físico (SMOLKA et al., 2000). Todos os ensaios enzimáticos foram feitos em duplicata.
Ensaios enzimáticos. A atividade da CS foi medida em meio contendo 0.1 mM de DTNB e 0.3 mM acetil CoA monitorando a queda da absorbância a 412 nm após a adição de 0.5 mM oxaloacetato (SRERE, 1969). A atividade da CS foi expressa em UI/mg tecido. A atividade da GR foi medida em tampão fosfato 0.2 M (pH 7.0), contendo ainda 2 mM EDTA, 0.1 mM NADPH e 0.75 mM DTNB. Para iniciar a reação, foi adicionado 1 mM de GSSG e a absorbância foi monitorada a 240 nm (SMITH et al.,1988). A atividade da GR foi expressa em UI/mg proteína. A atividade da CAT foi medida em tampão fosfato 50 mM (pH 7.0), monitorando a queda da absorbância a 240 nm durante 30 segundos após a adição de 10 mM de H2O2 (AEBI, 1984). A atividade da CAT foi expressa em UI/mg tecido.
Amostras de urina e ensaio de substâncias reativas ao ácido tiobarbitúrico (TBARs). Uma vez por semana, após a última sessão de treino, os animais eram colocados em gaiolas metabólicas por 24 horas. A urina era então coletada em toluene e armazenada a - 70oC até sua utilização para os ensaios de TBARs segundo YAGI (1987). Uma curva padrão de tetraetoxipropano foi utilizada como padrão para efetuar os cálculos da concentração de TBARs. A concentração de TBARs foi normalizada através da concentração de creatinina (JENKINS et al., 1992), que foi determinada utilizando um kit fotométrico Boehringer Mannheim específico.
Análise estatistica. Os dados estão expressos em média ±EP indicado por barras verticais nas figuras. Os grupos são compostos por oito animais (n=8). Foi utilizado o programa GraphPad Instat® (San Diego, CA) para aplicar o teste de ANOVA e o teste de Tukey como pós-teste, estabelecendo P <0.05 como limite confidencial.
ResultadosPerfil da intensidade ao longo do treinamento. A intensidade empregada semanalmente no treinamento foi inferida pela quantificação da concentração de lactate plasmático nos grupos experimentais. A figura 1 mostra que os protocolos de treinamento recrutaram predominantemente vias metabólicas de ressíntese de ATP diferentes. Estes dados mostram que a principal via recrutada no grupo CT foi a via oxidativa ou metabolismo aeróbico, indicando que este protocolo foi menos intenso na maior parte do tempo quando comparado ao protocolo de IT. O oposto foi observado no protocolo de IT, que durante quase todo o período de treinamento mostrou concentrações de lactato significativamente maiores comparadas aos grupos CT e SED, comportamento já esperado para um protocolo de exercício de alta intensidade. Além disso, observamos ainda que ambos os protocolos induziram sobrecarga progressiva na fase I e que essa fase foi mais intensa quando comparada à fase II.
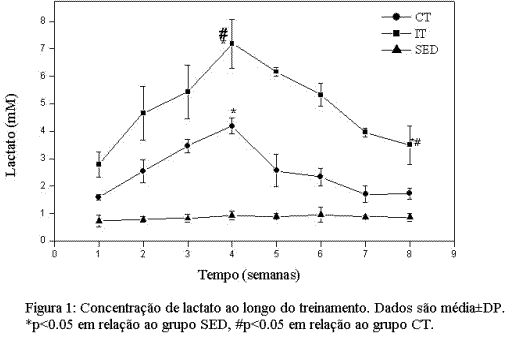
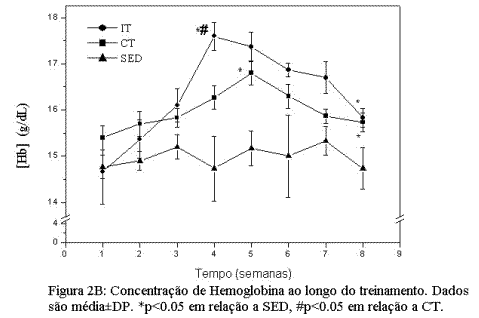
Adaptações induzidas pelos treinamentos de endurance e intervalado no metabolismo oxidativo.A figura 2A mostra a variação na atividade da CS, nos três grupos experimentais (SED, CT e IT), medida ao final das duas fases do treinamento. Podemos observar que, após a fase I, ambos os protocolos se mostraram eficientes em aumentar significativamente a atividade da CS. Ao final da fase II, no entanto, encontramos atividade significativamente maior da CS no grupo IT em relação aos grupos CT e SED. As alterações na concentração de Hb (Figura 2B) mostram um aumento significativo (p<0.05), atingindo seu pico máximo na quarta e quinta semanas de treinamento nos grupos CT e IT respectivamente, seguido pela estabilização em valores abaixo do atingido no pico máximo, ao final da fase II, embora se mantendo acima (p<0.05) dos valores encontrados no grupo SED.
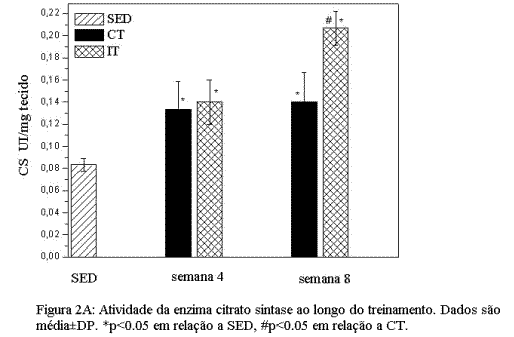
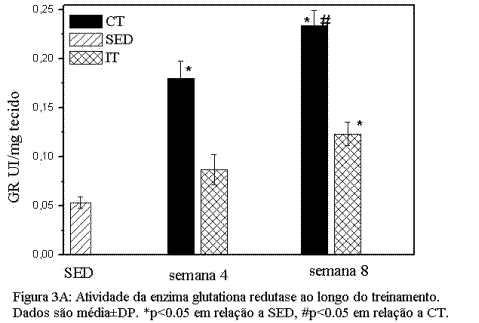
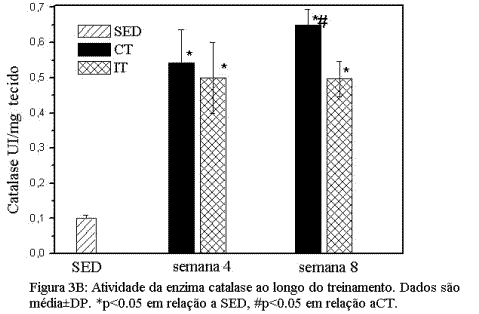
Adaptações induzidas pelo treinamento de endurance e intervalado nas enzimas do sistema antioxidante.As figuras 3 A e B mostram as adaptações induzidas nos biomarcadores do sistema enzimático de defesa antioxidante no músculo sóleo. A Figura 3 mostra que ambos os protocolos de treinamento induziram aumento significativo (p<0.05) nas enzimas GR e CAT quando comparadas ao grupo SED. No entanto, o grupo CT mostrou aumento mais efetivo das enzimas antioxidantes estudadas nas duas fases do treinamento.
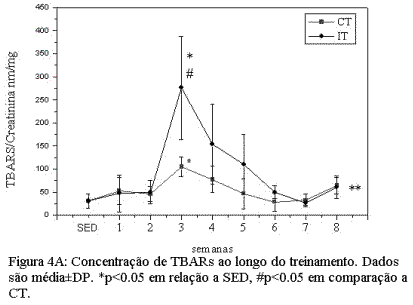
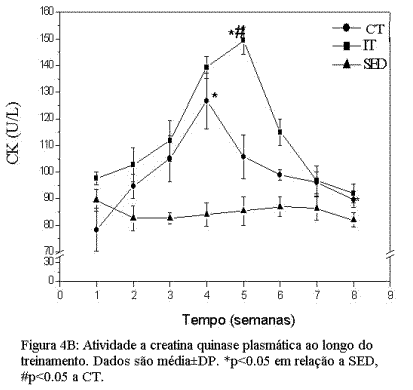
Alterações induzidas pelo treinamento de endurance e intervalado nos níveis de peroxidação lipídica e dano muscular.A figura 4A mostra as alterações induzidas pelos protocolos de CT e IT na concentração de TBARs e na atividade da CK plasmática (figura 4B). Nossos dados mostram que, para ambos grupos treinados, houve um aumento significativo na concentração urinária de TBARs, que atingiu seu pico máximo ao final da terceira semana de treinamento (Figura 4A). Este aumento foi particularmente maior (p<0.05) para o grupo IT em comparação ao grupo CT e SED. Após atingir seu pico máximo, os níveis de TBARs tenderam a cair para próximo dos valores do grupo SED ao final dos protocolos de CT e IT. A cinética da atividade da CK plasmática, ao longo de oito semanas de treinamento, mostrou comportamento similar à concentração de TBARs (figura 4A). Para esta variável, no entanto, o pico máximo de atividade ocorreu entre a quarta e quinta semana, nos grupos CT e IT respectivamente.
DiscussãoDados consistentes da literatura demonstram o aumento induzido pelo exercício físico nos níveis de estresse oxidativo (BRUIN et al., 1994; SJODIN et al., 1990; VINA et al., 2000; CANNON et al., 1990; TIIDUS, 1998; SMOLKA et al., 2000), fenômeno confirmado também por este estudo.
A questão se este processo irá levar a adaptação dos sistemas fisiológicos envolvidos (JACKSON, 1999; SMITH, 2000) ou ao dano oxidativo excessivo, e em última instância, lesões musculares (SJODIN et al.,1990; HESSEL et al., 2000; TIIDUS,1998), é de extrema importância na aplicação de programas de treinamento.
Estudos mostram que baixos níveis de marcadores de dano oxidativo se correlacionam com uma resposta adaptativa positiva e conseqüente melhora do condicionamento muscular ao esforço (JI,1999; RADAK et al.,1999). Algumas evidências demonstram, que a atividade física moderada induz aumento nas enzimas antioxidantes em músculo e também nas hemáceas (DUTHIE et al., 1990; POWERS et al., 1999), embora estudos anteriores sugiram que o nível de estresse oxidativo possa ser modulado tanto pela intensidade, como duração e ainda o tipo de atividade (SEN, 1995; SMOLKA et al., 2000).
Dados com relação aos efeitos de atividades intermitentes de alta intensidade sobre os níveis de estresse oxidativo e atividade das enzimas do metabolismo oxidativo e sistemas antioxidantes são escassos, permanecendo questionáveis as adaptações induzidas por este tipo de atividade (ESSIG & NOSEK, 1997; HALLIWELL & GUTTERIDGE, 1998).
Com o objetivo de estudar estas questões, neste trabalho, dividimos os protocolos de treinamento CT e IT em duas fases distintas (I e II) com intensidade e duração diferenciadas.
A fase I teve como característica maiores intensidades e menor volume em ambos os protocolos de treinamento. Em comparação aos dois grupos, ainda na fase I, IT foi de maior intensidade e menor volume em relação ao CT, conforme mostra a concentração de lactato na figura 1 e também à distância percorrida pelos dois grupos experimentais durante esta fase. Já na fase II a concentração de lactato nos mostra que a intensidade foi similar, enquanto o volume de esforço, medido através da distância percorrida, executado pelo grupo IT foi ainda maior que o grupo CT. Portanto, a análise das duas fases distintas dos protocolos de treinamento nos dá uma noção do efeito da intensidade (fase I) e do volume (fase II) nos marcadores estudados.
Neste sentido, nossos dados mostram que, para ambos os grupos de treinamento CT e IT, o aumento do volume e da intensidade empregados nos esforços induziram aumento nos níveis de estresse oxidativo, embora o protocolo de IT se mostrou mais eficiente como indutor de estresse. Tal fato ficou demonstrado quando observamos na fase I, onde a intensidade de esforço era mais alta, este protocolo induziu menor aumento na atividade das enzimas antioxidantes, acompanhado de maiores níveis de peroxidação lipídica e lesão muscular, mostrados pela concentração de TBARs na urina e atividade da CK plasmática.
Os altos níveis de lactato no grupo IT demonstram que a taxa de utilização de ATP excedeu a capacidade de sua ressíntese mitocondrial, embora o fluxo de oxigênio nas fibras musculares estivesse elevado devido ao aumento em sua capacidade de transporte e utilização como mostrados nas figuras 2A e 2B.
Nossos resultados confirmam ainda que, o estresse oxidativo causado pelo exercício de alta intensidade, ocorre na presença de oxigênio (SJODIN et al., 1990). Além disso, estes dados mostram que a alta disponibilidade de oxigênio em conjunto com alta capacidade oxidativa leva a uma intensa produção de EROs , superando a capacidade intracelular de detoxificação das mesmas (SMOLKA et al., 2000).
Tal fato contribui para um maior aumento nos níveis de estresse oxidativo no grupo IT quando comparado ao grupo CT, situação que parece ser importante no aumento da enzima mitocondrial CS (SMOLKA et al., 2000).
Estes dados estão em acordo com demais estudos, que provam ser o treinamento intervalado de alta intensidade um estímulo mais eficiente para aumentar a capacidade oxidativa intramuscular, incluindo a capacidade de oxidação de ácidos graxos, quando comparado ao treinamento de endurance (CHILIBECK et al., 1997; TERADA et al., 2004).
A liberação de proteínas musculares (i.e. CK), que normalmente são incapazes de atravessar a membrana celular, reflete uma mudança nas características de permeabilidade da mesma, permitindo a passagem de moléculas de grande peso molecular como as proteínas (ORTENBLAD et al., 1991).
Este fenômeno é normalmente associado pela literatura, como evidência de dano muscular (NOSAKA E NEWTON, 2002). É interessante destacar ainda que apenas o grupo IT mostrou correlação positiva (r = 0.92; P < 0.05) entre a concentração de TBARs, marcador de peroxidação lipídica e a atvidade de CK plasmática.
Estes dados, nos permitem inferir que a sobrecarga progressiva, principalmente associada ao aumento da intensidade de esforço, induz maior aumento nos níveis de estresse oxidativo devido à combinação de uma maior capacidade oxidativa em conjunto com aumento menos eminente na atividade das enzimas antioxidantes, o que levou o grupo IT neste estudo, a estar mais vulnerável a lesões musculares de origem oxidativa.
Nossa hipótese é, portanto, que o processo adaptativo deve estar em acordo com a demanda funcional das fibras musculares imposta pelo exercício físico. Em outras palavras, um protocolo de treinamento ideal não deve apenas aumentar a capacidade oxidativa muscular, mas também aumentar a capacidade de detoxificação de EROs produzidas durante o esforço.
O aumento dessa capacidade seria atingido principalmente através do aumento na atividade das enzimas antioxidantes e dos demais mecanismos de proteção intracelular, tais como, a expressão das proteínas de estresse (SMOLKA et al., 2000).
Nossos dados ainda mostraram que os marcadores escolhidos são sensíveis em demonstrar o processo adaptativo tanto no músculo sóleo quanto nas hemáceas, fato que pode ser observado ao final do treinamento.
Concluindo, nossos dados permitem inferir que esforços executados em alta intensidade são mais eficientes em induzir aumento na capacidade oxidativa muscular, enquanto atividades de menor intensidade e maior volume parecem atuar de forma mais eficiente no aumento do sistema antioxidante enzimático e, ainda, que o acompanhamento dos marcadores estudados neste trabalho pode demonstrar o nível de estresse imposto pelo treinamento físico, o que se pode constituir numa ferramenta importante para garantir apenas o efeito adaptativo do treinamento evitando, assim, lesões musculares mais graves.
AgradecimentosOs autores agradecem à Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP) E CNPq pelo suporte financeiro para este estudo.
Abreviações utilizadasEDTA: Ácido etilenodiaminotetracético
GSSG: Glutationa dissulfeto
MOPS: Ácido 4-Morpholinopropanosulfônico
NADPH: Adenina dinucleotídeo fosfato
PMSF: Fenil metil sulfonil fenazina
DTNB: Ácido 5,5'-Ditiobis-2-nitrobenzóico
Referências bibliográficas
AEBI, H. Catalase in vitro. Methods Enzymology v.105, p.121-126, 1984.
ALESSIO, H. M. Exercise-induced oxidative stress. Medicine and Science in Sports and Exercise. v. 25,n.2, p. 218-224, 1993.
ALESSIO, H.M., et al. Generation of reactive oxygen species after exhaustive aerobic and isometric exercise. Medicine and Science in Sports and Exercise. v.32, n. 9, p.1576-1581, 2000.
Bishop, D. Evaluation of the Accusport lactate analyser. International Journal of Sports Medicine, v.22, n. 7, p.525-30, 2001.
BRUIN, G. et al. Adaptation and overtraining in horses subjected to increasing training loads. Adaptation and overtraining in horses subjected to increasing training loads. Journal of Applied Physiology, v.76, n. 5, p. 1908-1913, 1994.
CANNON, J.G. et al. Acute phase response in exercise. II. Associations between vitamin E, cytokines, and muscle proteolysis. American Journal of Physiology, v.260, n. 6 pt. 2, p. R1235-R1240, 1991.
CANNON, J.G. et al. Acute phase response in exercise: interaction of age and vitamin E on neutrophils and muscle enzyme release. American Journal of Physiology, v. 259, n. 6 pt 2, p. R1214-R1219, 1990.
CHILIBECK, P.D. et al. Higher mitochondrial fatty acid oxidation following intermittent versus continuous endurance exercise training. Canadian Journal of Physiology and Pharmacology, v. 76, n. 9, p. 891-894, 1997.
CLANTON, T.L. et al. Oxidants and skeletal muscle function; Physiologic and pathophysiologic implications. Proceedings of the Society of Experimental Biology Medicine, v. 222, p. 253-262, 1999.
CRISWELL, D. et al. High intensity training-induced changes in skeletal muscle antioxidant enzymes activity. Medicine and Science in Sports and Exercise, v. 25, n. 10, p. 1135-1140, 1993.
DAVIES, K.J. et al. Free radicals and tissue damage produced by exercise. Biochemical and Biophysical Research Communications, v. 107, n. 4, p. 1198-1205, 1982.
DILLARD, C.J. et al. Effects of exercise, vitamin E, and ozone on pulmonary function and lipid peroxidation. Journal of Applied Physiology, v. 45, n. 6, p. 927-932, 1978.
DUTHIE, G.G. et al. Blood antioxidant status and erytrocyte lipid peroxidation following distance running. Archives of Biochemistry Biophysics. V. 282, n. 1, p. 78-93, 1990.
ESSIG, D.A. and NOSEK, T.M. Muscle fatigue and induction of stress protein genes: a dual function of reactive oxygen species? Canadian Journal of Applied Physiology, v. 22, n. 5, p. 409-428, 1997.
Fell, J.W. Evaluation of the Accusport Lactate Analyser. International Journal of Sports Medicine, v.19, n.3, p.199-204, 1998.
FIELDING, R.A. et al. Acute phase response in exercise. III. Neutrophil and IL-1 beta accumulation in skeletal muscle. American Journal of Physiology, v. 265, n. 1 pt 2, p. R166-R172, 1993.
GLEESON, M. Interleukins and exercise. Journal of Physiology, v. 15, n. 1 pt 1, p. 529, 2000.
HALLIWELL, B. and GUTTERIDGE. J.M.C. Oxidative Stress: adaptation, damage, repair and death. Oxford: Oxford University Press, 1998.
HESSEL, E. et al. Oxygen radical generation of neutrophils: a reason for oxidative Oxygen radical generation of neutrophils: a reason for oxidative Clinical Chemical Acta v. 298, n. 1-2, p.145-156, 2000.
JACKSON, M.J. An overview of methods for assessment of free radical activity in biology. Proceedings of the Nutritional Society, v.58, n. 4, p.673-676, 1999.
JI, L.L. and LEICHTWEISS, S. Exercise and oxidative stress: Sources of free radicals and their impact on antioxidant systems. Age, v. 20, p. 91-106, 1997.
JI, L.L. Exercise and oxidative stress in exercise. Proceedings of the Society of Experimental Biology Medicine, v.222, n. 3, p. 283-292, 1999.
JENKINS, R.R. et al. Influence of exercise on clearance of oxidant stress products and loosely bound iron. Medicine and Science in Sports and Exercise, v.25, n. 2, p.213-217, 1993.
KUIPERS, H. Training and overtraining: an introduction. Medicine and Science in Sports and Exercise, v.30, n. 7, p.1137-1139, 1998.
LIU, J.K. et al. Chronically and acutely exercised rats: biomarkers of oxidative stress and endogenous antioxidants. Journal of Applied Physiology, v.89, p.21-28, 2000.
NOSAKA, K. and NEWTON, M. Concentric or eccentric training effect on eccentric exercise-induced muscle damage. Medicine and Science in Sports and Exercise, v.34, n.1, p.63-69, 2002.
ORTENBLAD, N. et al. Antioxidant status and lipid peroxidation after short-term maximal exercise in trained and untrained. American Journal of Physiology, v.260, p. R1235-R1240, 1991.
POWERS, S.K. and LENNON, S.L. Analysis of cellular responses to free radicals: focus on exercise and skeletal muscle. Proceedings of the Nutritional Society, v.58, n. 4, p. 1025-1033, 1999.
POWERS, S.K. et al. Exercise training-induced alterations in skeletal muscle antioxidant capacity: a brief review. Medicine and Science in Sports and Exercise, v.31, n. 7, p.987-997, 1999.
RADÁK, Z. et al. The effect of exercise training on oxidative damage of lipids, proteins and DNA in rat skeletal muscle: evidence for beneficial outcomes. Free Radicals in Biology and Medicine, v. 27, n. 1-2, p. 69-74, 1999.
REID, M.B. et al. Reactive oxygen in skeletal muscle. II. Extracellular release of free radicals. Journal of Applied Physiology, v. 73, n. 5, p. 1805-1809, 1992.
SEN, C.K. Oxidants and antioxidants in exercise. Oxidants and antioxidants in exercise. Journal of Applied Physiology, v.79, n.3, p.675-686, 1995.
SJODIN, B. et al. Biochemical mechanisms for oxygen free radical formation during exercise. Sports Medicine v.10, n. 4, p. 236-254, 1990.
SMITH, I.K. et al. Assay of glutathione reductase in crude tissue homogenates using 5,5'-dithio-bis(2-nitrobenzoic acid). Analytical Biochemistry, v. 175, n. 2, p. 408-413, 1988.
SMITH, L.L. Cytokine hypothesis of overtraining: a physiological adaptation to excessive stress? Cytokine hypothesis of overtraining: a physiological adaptation to excessive stress? Medicine and Science in Sports and Exercise, v.32, n. 2, p.317-331, 2000.
SMOLKA, M.B. et al. HSP72 as a complementary protection against exercise induced oxidative stress in the soleus muscle of rats. American Journal of Physiology, v.279, n. 5, p. R1539-R1545, 2000.
SRERE, P.A. Citrate synthase. Methods Enzymology, v.13, p.3-11, 1969.
TERADA, S. et al. Effect of high-intensity intermittent swimming training on fatty acid oxidation enzyme activity in rat skeletal muscle. Japanese Journal of Physiology, v. 54, n.1, p.47-52, 2004.
TIIDUS, P.M. Radical species in inflammation and overtraining. Canadian Journal of Physiology and Pharmacology, v. 76, n. 5, p. 533-538, 1998.
VINA, J. et al. Mechanism of free radical production in exhaustive exercise in humans and rats; role of xanthine oxidase and protection by allopurinol. IUBMB Life, v. 49, n. 6, p. 539-544, 2000.
VIRU, A. The mechanism of training effects: a hypothesis. International Journal of Sports Medicine, v. 5, n. 5, p. 219-227, 1984.
YAGI, K. Lipid peroxides and human diseases. Chemistry and Physics of Lipids, v.45, n. 2-4, p. 337-351, 1987.
ZERBETTO, E. et al. Quantification of muscle mitochondrial oxidative phosphorylation enzymes via histochemical staining of blue native polyacrylamide gels. Electrophoresis, v. 18, n. 11, p. 2059-2064, 1997.
Legendas

| |
|
|---|---|
|
revista
digital · Año 11 · N° 99 | Buenos Aires, Agosto 2006 |
|