
|
|||
|---|---|---|---|
|
|
Efectos de los suplementos de creatina sobre el rendimiento físico Effects of the creatine supplements on the physical performance |
|
|
|
Licenciado en Educación Física Profesor de Educación Física Preparador Físico |
Raúl Gonzalo Prieto raulgonzalo@yahoo.es (España) |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 10 - N° 69 - Febrero de 2004 |
|
|
1 / 1
Introducción
1.1. Concepto de creatina y fosfocreatina
La creatina (C4H9O2N3), también denominada ácido guanidinacético, metilglicociamina, N-metil-N-guanilglicina o ácido (alfa-metilguanido) acético; tiene como fórmula química la representada en la FIGURA 1:
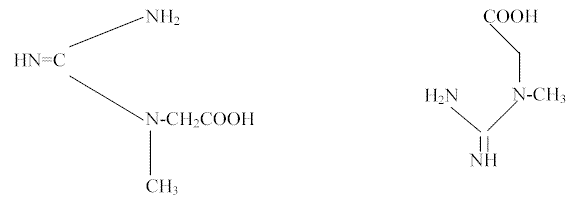
Figura 1Fue descubierta, en 1835, por Chevreul, en el caldo de la carne y estudiada detenidamente por Liebig en 1847. Es un aminoácido muy especial pues no es uno de los del grupo alfa, que entran en la composición de las proteínas; sino un producto final del metabolismo de la glicina, de la arginina y de la metionina, que se encuentra predominantemente en el músculo de todos los mamíferos, aves, anfibios y peces y también en el cerebro, sangre y otras partes del organismo de éstos.
Forma parte de las llamadas sustancias extractivas del músculo que comprenden una serie de sustancias orgánicas: ácido inosínico, carnosina (0,2 %), creatina (0,4 %), carnitina, inosita, etc.; de bajo peso molecular, que se pueden separar del músculo por ebullición con agua.
La fosfocreatina (C4H9PO5N3) también denominada ácido creatin-fosfórico y creatin-fosfato (Ver FIGURA 2) fue descubierta por P. Eggleton y G. P. Eggleton y denominada fosfágeno. Fiske y Subbarow demostraron que contenía creatina y finalmente Meyerhof y Lohmann comprobaron que se compone de 1 mol de creatina y 1 mol de ácido fosfórico.
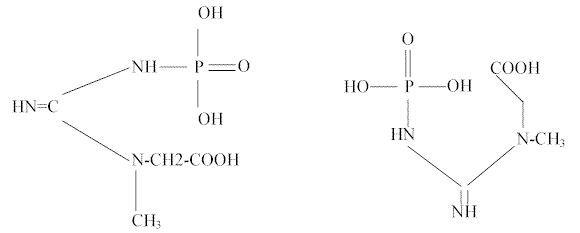
Figura 2Trabajos posteriores demostraron que la fosfocreatina se descomponía, para formar creatina, cuando se activaba el músculo envenenado con iodoacetato y que el trabajo realizado por éste, era equivalente a la descomposición del fosfágeno.
1.2. La Fosfocreatina (PCr) como depósito de energíaLa contracción muscular depende de la energía proporcionada por el ATP. La mayor parte de esta energía se necesita para mover el mecanismo de cremallera por virtud del cual los puentes cruzados tiran de los filamentos de actina, pero se necesitan cantidades pequeñas para bombear calcio del sarcoplasma (que contiene además fosfocreatina, proteínas libres, gotitas de lípidos y numerosísimas mitocondrias dispuestas a lo largo de las miofibrillas), hacia el interior del retículo sarcoplásmico, y bombear iones de sodio y potasio, a través de la membrana de la fibra muscular para conservar un medio iónico adecuado que permita la propagación de potenciales de acción.
Sin embargo, la cantidad de ATP presente en la fibra muscular sólo basta para conservar la contracción plena durante menos de un segundo. Afortunadamente, después de que el ATP se ha roto en ADP, este último se refosforila para formar nuevo ATP en el plazo de una fracción de segundo, gracias a la intervención del fosfato de creatina.
Lundsgaard, con motivo de sus observaciones sobre la contracción aláctica, desarrolló su hipótesis de que la reacción primaria que suministra la energía para la contracción no es la formación de ácido láctico, sino el desdoblamiento de la fosfocreatina. Este punto de vista se confirma gracias a los posteriores trabajos de Meyerhof y su escuela.
Hemos de tener en cuenta que en el metabolismo anaeróbico, podemos obtener ATP a través de tres vías:
Mediante la reacción catalizada por la enzima adenilatoquinasa, en la que se forman ATP y AMP (Ver FIGURA 3).
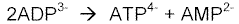
Figura 3
Mediante la glucólisis anaeróbica, en la que el glucógeno muscular se degrada a ácido láctico, a través de unos cuantos poros enzimáticos, con ATP sintetizado en el proceso por fosforilación de ADP a nivel de sustrato.
Mediante la reacción catalizada por la enzima creatin-quinasa, donde la fosfocreatina (PC), puede desdoblarse para formar ATP y creatina (Cr) (Ver Figura 4).
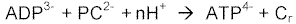
Figura 4El fosfato de creatina o fosfocreatina lleva un enlace fosfático rico en energía similar a los del ATP, aunque a diferencia de éste, no puede servir de intermediario en el transporte de energía de los alimentos a los sistemas funcionales celulares.
El ATP constituye la fuente energética inmediata para el trabajo muscular pero su concentración es solamente de 5 mM/kg de tejido muscular. Sin embargo, la concentración de la fosfocreatina es de 20 mM/kg de tejido (20 a 30 mol/ g de tejido, proporcionando una reserva de grupos fosforilo de alta energía, muy importante en la regeneración del ATP (mantiene el sistema del adenilato del músculo regulado, en un estado de "carga energética" elevada).
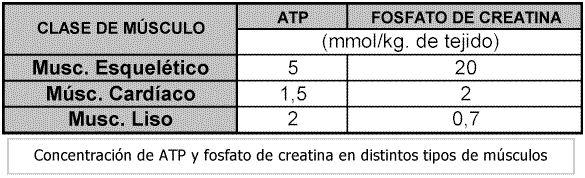
Figura 5 (Mohler, J., 1984)Además encierra unas 8.500 cal/mol, en condiciones estándar, ó 9.500 cal/mol, en los del cuerpo. (38ºC y bajas concentraciones de los reactivos).
Esta cifra es vecina de las 8.000 cal/ mol, que corresponden a cada uno de los dos enlaces fosfato del ATP. Este desnivel energético entre los enlaces de las dos sustancias provoca que la reacción sea más fácil en el sentido fosfato de creatina
ATP.
Por tanto, cuando comienza la utilización de ATP, éste se sintetiza de nuevos a expensas de la energía almacenada en el fosfato de creatina (durante los primeros segundos). Este efecto conserva la concentración de ATP casi constante, siempre y cuando quede un poco de creatina fosfato. Por esta razón, podemos realmente denominar el sistema ATP-creatina fosfato un sistema amortiguador del ATP.
De hecho, es fácil comprender la importancia de conservar casi constante la concentración de ATP, porque la velocidad de casi todas las reacciones del organismo dependen de esta constancia. (Ver FIGURA 6).
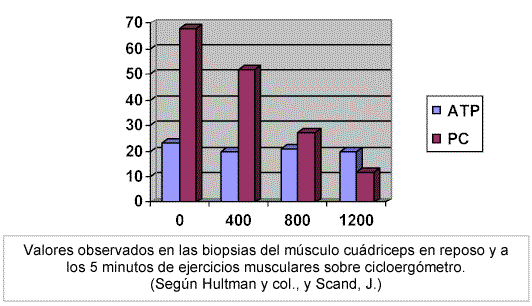
Figura 6 (Monod, H. y Flandrois, R., 1986)Aún así, el número de enlaces fosfato contenidos como tales, en el músculo bien como ATP, bien almacenados como fosfocreatina, es muy limitado (por la baja concentración intramuscular de ambos metabolitos) y no son posibles más allá de 50 a 100 concentraciones antes de que se produzca la depleción total.
En una contracción intensa, ello significa aproximadamente una duración entre 5 y 8 segundos, aunque este período sea muy variable en función de las características del músculo y del nivel de entrenamiento del mismo, entre otras causas.
A pesar de que en la actividad motora usual, el papel cuantitativo de estas reservas de fosfágeno es escaso, porque la duración de los períodos contráctiles es por lo general mucho más prolongada, estos enlaces fosfato son decisivos para permitir iniciar la contracción y aún mantenerla el tiempo necesario para que terminen de ser completadas el conjunto de reacciones oxidativas de los sustratos disponibles, absolutamente necesarias para la obtención de la energía química que permita la puesta en marcha de la actividad fosforilativa, generadora de nuevos enlaces fosfato.
Se necesita ATP para la refosforilación de la creatina a fosfato de creatina. Éste cede a la creatina uno de sus restos fosfato, según la reacción de Lohman (ver FIGURA 7).
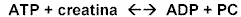
Figura 7Con este proceso, la creatin-quinasa ectomitocondrial (ligada exteriormente a las mitocondrias), podría desempeñar el papel más importante, mientras que la enzima citoplasmática tendría más importancia en la reacción que transcurre en el sentido contrario.
Como se puede suponer, la reacción de la creatin-quinasa, para la producción de ATP, entraña la eliminación de "n" iones H+ por cada fosfocreatina utilizada. (El número de iones "n" se refiere a los realmente consumidos en función del pH). A medida que este desciende, la reacción regulada por la creatin-quinasa capta relativamente más iones H+ y provoca una alcalinización relativa en el comienzo del ejercicio de acuerdo con el proceso predominante de producción de ATP.
Después, a medida que entra en juego la glucólisis anaerobia, a poco de empezado el ejercicio y cuando aumenta el contenido de ATP de la fibras hasta alcanzar niveles suficientes para invertir el sentido de la reacción, se acopla, en lo funcional, a la reacción regulada por la creatin-quinasa, como muestra la FIGURA 8:
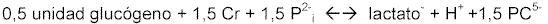
Figura 8 (Bove, A. A. y Lowenthal, D. T., 1997)Esto revierte efectivamente el proceso de alcalinización y pasa a otro de formación neta de iones H+ que tiende a una relativa acidificación del músculo en actividad.
La degradación anaeróbica del glucógeno suministra, pues, la energía necesaria para la regeneración constante del fosfato de creatina. Naturalmente, todos los procesos oxidativos que suministran ATP pueden intervenir también en la regeneración de la fosfocreatina. La importancia de la glucólisis, consiste pues en que trabaja también en condiciones anaeróbicas.
Aunque es posible que todo este proceso tenga lugar durante la contracción (si su baja intensidad lo permite), se completa, por lo general, durante la relajación muscular; por lo tanto, no se recuperan completamente los niveles de fosfocreatina iniciales hasta que se alcanza la condición de reposo. Esta recuperación es rápida, no superior a 1 ó 2 minutos, aunque se ve modificada por diversos factores: fatiga, formación de lactato, etc.
Entonces, como hemos visto, el ATP refosforila, en última instancia, la creatina a fosfocreatina y parece claro que esto se produce porque el fosfato de creatina es muy útil como reserva adicional de compuestos de alta energía, que serviría en los episodios repentinos de intensa actividad.
Sin embargo, ¿no podría lograrse sólo con mantener un depósito más grande de ATP, puesto que normalmente las mitocondrias trabajan muy por debajo de su capacidad máxima? La explicación de todo ello puede venir de la posibilidad de que la fosfocreatina participe en un sistema de recambio, para transferir la energía del ATP, desde las mitocondrias, donde se forma, hasta el citoplasma, donde se utiliza (como en la contracción muscular). Esto sería beneficioso pues la membrana mitocondrial no es del todo permeable al ATP.
De acuerdo con este plan, a medida que el ATP se utiliza en las actividades que se desarrollan en el citoplasma, se repone inmediatamente por medio del fosfato de creatina, la creatina resultante difunde a la superficie de la mitocondria: en la pared de la mitocondria hay una forma fija de la enzima creatina-fosfoquinasa, que transfiere un grupo fosfato de alta energía, a través de la membrana, desde el ATP mitocondrial hasta la creatina citoplasmática, de modo que se repone la fosfocreatina citoplasmática y queda, una vez más, disponible para mantener una alta concentración de ATP en el citoplasma, donde se le necesita para diversas actividades celulares.
El ADP que se forma en la mitocondria con esta reacción se refosforila enseguida de la manera habitual, por medio de la cadena respiratoria. El resultado es que la energía generada en la mitocondria queda rápidamente a disposición del citoplasma, donde se necesita para realizar trabajo. Esta hipótesis concuerda con el hecho de que sólo existen grandes concentraciones de fosfocreatina en tres órganos de requerimiento energético sea en forma constante (encéfalo, corazón), u ocasional (músculo esquelético). Además, en la membrana mitocondrial hay otros transportadores que facilitan el traslado de ATP desde las mitocondrias hasta el citoplasma.
Por último, cabe reseñar que la creatina no enzimática, después de perder su contenido de H20 se transforma en creatinina, para ser eliminada por la orina.
Efectos de la suplementación con creatina en el rendimiento físico.En 1926 aparecen datos que indican que las reservas intramusculares de creatina pueden aumentarse ingiriendo cantidades mayores de las normales. Se postuló que la suplementación con creatina mejora la actividad de los deportistas, aumentando la fuerza muscular, incrementando la energía muscular (mayor energía disponible por unidad de tiempo) y acelerando el tiempo de recuperación entre ejercicios intensos y de corta duración. Uno de los primeros estudios, el de Sipila y col. (1981) (citado por Rico Sanz, J. y col., 1997) concluye un aumento del peso corporal de un 10 % y un aumento de un 32 % en el diámetro de las fibras musculares Tipo II.
Posteriormente autores como Francaux y col. (2000) y Dorado García y col. (1997) coincidieron en que con el aporte suplementario de creatina, la cantidad total de este compuesto en el músculo esquelético puede aumentar por encima del 30 % y la cantidad de fosfocreatina por encima del 20 % y mejorar el rendimiento en esfuerzos de muy corta duración. Posiblemente esto se deba a que la concentración elevada de fosfocreatina antes de iniciarse el ejercicio podría reducir la velocidad de la glucólisis al menos por tres mecanismos: inhibición de la fosfofructoquinasa, reducción potencial de la producción de ADP y AMP y aumento de la capacidad tampón del músculo (hasta un 7 %).
Balsom y cols. (1993) investigaron si la suplementación con creatina retrasa la aparición de la fatiga, durante series de elevada intensidad y corta duración, y estudiaron cualquier modificación asociada en el metabolismo energético. Este estudio demostró que el aumento del contenido total de creatina en el músculo por administración de las mismas puede retrasar la aparición de la fatiga durante series repetidas de ejercicios de gran intensidad.
Además no se han descrito hasta el presente efectos colaterales asociados a la ingesta de grandes cantidades de creatina, lo que muestra una excelente relación beneficio-riesgo (Paús, V. y col., 1998).
En nuestro país una de las investigaciones más interesantes realizadas en este ámbito, ha sido la iniciada en 1993, en el C.A.R. (Centro de Alto Rendimiento) de Sant Cugat (Ruíz, A., 1995).
Tras el éxito de L. Christie y de los velocistas ingleses, en los JJ.00. de Barcelona, se supo que tomaban creatina y que esta sustancia era responsable, en parte, de sus grandes resultados.
Por ello, la Unidad de Investigación del C.A.R. y el entrenador de los atletas de velocidad del C.A.R., Alex Codina, plantearon realizar un estudio para ver si era útil o no la ingesta de creatina en los velocistas.
En la temporada de 1993, se diseñó un test para ver si la suplementación con creatina incrementaba el rendimiento en las pruebas explosivas. También realizaron un profundo estudio de toda la bibliografía publicada sobre la creatina y, así, determinaron la cantidad de creatina a tomar y el tiempo de ingesta.
Su primera experiencia fue administrar 10 g diarios durante tres semanas. Con una serie de test de entrenamiento y con el seguimiento del mismos intuyeron que el efecto de la creatina era positivo. Esta primera toma de contacto resultó satisfactoria pero, por problemas logísticos, y de calendario los estudios no se terminaron.
En la temporada de 1994, se planteó realizar los estudios tal y como se habían previsto realizar desde el principio pudiendo realizar las resonancia magnéticas adecuadas, que permitieron comprobar los resultados que provocan y en los atletas, la ingesta de creatina y el entrenamiento.
Se ajustaron las dosis de suplementación (que fueron de 21 g durante seis días), en tres períodos de entrenamiento. Se efectuó una resonancia magnética antes de la tomas, otra después de tomar la creatina durante seis días y, por fin otra pasados 15 días después de la toma. En ellas, se observó que existía un incremento de los depósitos de fosfocreatina en el músculo y que, al cabo de los 15 días de suspenderse la toma, los depósitos eran, aún, superiores a los iniciales.
Posteriormente, diseñaron una batería de test para determinar la utilización de estos depósitos y si, además, esto influía en el rendimiento del deportista. Para ello, dispusieron de un ergómetro que les permitía realizar ejercicios cíclicos en el interior del túnel de resonancia. Dicha batería consistió en:
Un test de 6 repeticiones de 10 segundos, a máxima intensidad, con recuperación de tres minutos.
Otro de 2 repeticiones de 30 segundos, con 8 minutos de recuperación.
Se efectuó una resonancia en reposo y luego otra con los ejercicios de la batería de test. Posteriormente se realizó una semana de suplementación (21 g durante 6 días), y se volvieron a realizar las resonancias de la batería de test. Durante los tiempos de recuperación, también se realizaron resonancias para determinar el comportamiento de la fosfocreatina y su recuperación además del pH muscular que permitió cuantificar la acidez del músculo.
Realizados los estudios descritos llegaron a las siguientes conclusiones:
El aumento de los depósitos de fosfocreatina produce un efecto tampón que reduce la acidez del músculo por lo que es posible incrementar la carga de entrenamiento, permitiendo realizar más repeticiones a la misma intensidad con recuperaciones más cortas. También, se puede incrementar la intensidad de trabajo, manteniendo la misma recuperación.
Lo anteriormente mencionado justifica la posibilidad de incrementar los entrenamientos y sus cargas, reduciendo los tiempos de recuperación; aunque exige aumentar los días de descanso en los microciclos de entrenamiento, pues, aunque energéticamente el músculo se recuperaba fácilmente de la fatiga, neuromuscularmente no sucedía lo mismo.
Sin embargo, podemos encontrar otros autores como Cooke y Odland (citados por Rico Sanz, J. y cols., 1997)) que no observaron mejoría en protocolos de ejercicios intensos después de la suplementación.
Paús y cols. (1998) realizaron un ensayo con 8 futbolistas profesionales del primer equipo del Club Gimnasia y Esgrima La Plata de Argentina. El análisis de los resultados permitió concluir que los datos obtenidos no fueron estadísticamente relevantes para asegurar los beneficios de la administración de la creatina. Se pudo comprobar la falta de toxicidad y el aumento de masa corporal total.
Cooke, y cols. (1995) pretendieron determinar los efectos de una suplementación (vía oral), con creatina, sobre el rendimiento físico durante una prueba, en cicloergómetro, de alta intensidad y corta duración (tipo "sprint").
Los resultados obtenidos sugirieron que, en varones sanos no entrenados, la administración oral (a modo de "sobrecarga"), no contribuye a mejorar el rendimiento (expresado como potencia o resistencia a la fatiga), durante un ejercicio continuo de alta intensidad.
En cuanto a los efectos que puede producir en ejercicios prolongados, Spriet (citado por Dorado García, C. y col., 1997) y Balsom y cols. (1993) (citado por Rico Sanz, J. y col., 1997) consideraron que la suplementación oral con creatina no mejora el rendimiento ni el V02, en un ejercicio continuo y prolongado y que, por tanto, los efectos ergogénicos de la creatina están restringidos fundamentalmente a ejercicios de corta duración y gran intensidad.
Después de analizar parte de la numerosa literatura existente, se observa que la gran mayoría de los investigadores coincide en señalar que la suplementación correcta y planificada de creatina acompañada de un adecuado entrenamiento eleva el rendimiento en pruebas de elevada intensidad y corta duración (Sipila y col., 1981; citado por Rico Sanz, J. y col., 1997; Francaux, M. y col., 2000; Dorado García, C. y col., 1997; Ruíz, A. y col., 1995) pero no en actividades en las que las reservas de fosfágenos no es determinante (actividades de larga duración) (Spriet, L. L.; citado por Dorado García, C. y col., 1997; Balsom y cols., 1993; citado por Rico Sanz, J. y col., 1997).
Además, parece ser que retrasa la aparición de la fatiga durante series repetidas de ejercicios de gran intensidad (Balsom y cols., 1993).
Aunque no se han encontrado efectos secundarios con pautas de suplementación adecuadas (Paús, V. y col., 1998), Jorge Candel, jefe de los Servicios Médicos del Valencia C.F., comentó, durante su participación en el VII Congreso Nacional de Medicina Deportiva (en el que expuso los resultados de una investigación sobre el efecto del uso de la creatina como ayuda ergonómica para mejorar el rendimiento en los deportistas) que "no hay que olvidar las posibles alteraciones gástricas que producen dosis elevadas".
Debe realizarse la suplementación con un seguimiento médico, con el fin de evitar malas interpretaciones que pudieran surgir a raíz del incremento de la creatinina plasmática provocado por la ingesta de creatina y que podría asociarse a una alteración renal.
Bibliografía
Balsom, P. D. y col. (1994). La suplementación con creatina per se no mejora el rendimiento en ejercicios de resistencia. Actualizaciones en Fisiología del Ejercicio. 2 (2): 72.
Balsom, P. D. y col. (1995). Suplementación de creatina y ejercicio intermitente dinámico de elevada intensidad. Actualizaciones en Fisiología del Ejercicio. 3 (1): 211.
Bove, A. A.; Lowenthal, D. J. (1997). Medicina del ejercicio. Editorial El Ateneo. Buenos Aires.
Cooke, S. y col. (1995). Efectos de la ingesta de creatina sobre la potencia y fatiga durante un ejercicio en cicloergómetro. Actualizaciones en Fisiología del Ejercicio. 3 (1): 214.
Dorado García, C. y col. (1997). Efectos de la administración de suplementos de creatina sobre el rendimiento. Archivos de Medicina del Deporte. FEMEDE. 14 (59): 213-220.
Francaux, M. y col. (2000). Effect of exogenous creatine supplementation on muscle PCr metabolism. International Journal of Sports Medicine 21 (2): 139-145.
Mohler, J. (1984). Bioquímica. Editorial Pirámide. Madrid.
Monod, H.; Flandrois, R. (1986). Manual de Fisiología del Deporte. Editorial Masson. Barcelona.
Nosaka, K.; Clarkson, P. M. (1996). Variability in Serum Creatine Kinase Response After Eccentric Exercise of the Elbow Flexors. International Journal of Sports Medicine. 17 (2): 120-126.
Paús, V. y col. (1998). Administración de creatina monohidrato como suplemento energético en jugadores de fútbol profesional. Rev. de la Asociación Argentina de Traumatología del Deporte. 5 (5): (Extraído de la Revista Digital de Educación Física y Deportes en http://www.efdeportes.com).
Rico Sanz, J. (1997). Efectos de suplementación de creatina en el metabolismo muscular y energético. Archivos de Medicina del Deporte. FEMEDE. 14 (61): 391-396.
Ruiz, A. (1995). La Investigación en el CAR sigue en marcha. Revista CAR News. 10: 12-13.
| |
|
|---|---|
|
revista
digital · Año 10 · N° 69 | Buenos Aires, Febrero 2004 |
|