
|
|||
|---|---|---|---|
|
|
Perfil metabólico durante o exercício físico: influência da intensidade e da duração do esforço sobre a utilização de substratos energéticos Metabolic profile during physical exercise: influence of effort intensity and duration on the utilization of energetic substrates |
|
|
|
Doutorando em Ciências da Motricidade (Biodinâmica da Motricidade Humana) Universidade Estadual Paulista - UNESP. Rio Claro - SP Instituto Latinoamericano de Actividad Física Terapêutica (ILAFIT) |
Gustavo Puggina Rogatto grogatto@hotmail.com (Brasil) |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 8 - N° 54 - Noviembre de 2002 |
|
|
1 / 1
Introdução
O exercício físico é uma condição onde ocorre um aumento da demanda energética do organismo visando a manutenção da atividade muscular. A energia derivada dos nutrientes ingeridos na alimentação tem fundamental importância para o fornecimento de energia química, contribuindo com a manutenção do trabalho muscular a partir da geração de adenosina trifosfato (ATP) (WILMORE & COSTILL, 1994; McARDLE, KATCH & KATCH, 1992; FOX, BOWERS & FOSS, 1991).
Dentre os vários sistemas envolvidos no fornecimento energético para ressíntese de ATP, podemos destacar o papel das reservas de substratos energéticos que, por diferentes vias de fornecimento de energia, contribuem para a constante homeostase energética (WILMORE & COSTILL, 1994).
A intensidade e/ou a duração do esforço, bem como o estado inicial das reservas de substratos energéticos e o nível de treinamento do atleta podem interferir sobre a predominância na ativação de uma ou de outra via metabólica, indicando maior utilização de um determinado substrato energético. Assim, os fosfatos de alta energia, os estoques de glicogênio muscular e hepático, e os lipídeos estocados nos adipócitos podem contribuir com maior ou menor magnitude com a geração de energia durante o exercício (WILMORE & COSTILL, 1994).
Atividades realizadas por um longo período de tempo podem apresentar um equilíbrio (“steady-state”) entre a capacidade de geração de energia e a demanda decorrente da atividade muscular. Contudo, nos momentos iniciais do esforço e em exercícios severos, a ativação das reservas de substratos energéticos torna-se fundamental para o atendimento da maior exigência metabólica. Desta forma, o funcionamento e/ou a ativação destas vias de fornecimento de energia tem como objetivo fornecer uma quantidade adequada de nutrientes para o desempenho da atividade muscular (WILMORE & COSTILL, 1994; McARDLE, KATCH & KATCH, 1992; FOX, BOWERS & FOSS, 1991).
Metabolismo e exercícioAs reservas de substratos energéticos dos diferentes tecidos têm fundamental papel na manutenção da homeostase orgânica uma vez que a impossibilidade de armazenamento de grandes quantidades de ATP nas células gera a contínua necessidade de sua ressíntese. As vias geradoras de energia pela utilização das reservas de substratos teciduais podem depender ou não da presença de oxigênio para a ocorrência de suas reações, sendo que a predominância destas vias metabólicas tem íntima relação com a duração e intensidade da atividade. O sistema de fornecimento de energia dependente de oxigênio (aeróbio) produz ATP continuamente, porém sua velocidade de produção é baixa. Nos momentos iniciais, devido ao aumento abrupto da necessidade energética, a ativação da via anaeróbia tem grande participação no fornecimento energético, em função do déficit inicial de oxigênio. Nesta fase inicial de transição o composto de alta energia creatina-fosfato (CP) é o principal responsável pelo desempenho do trabalho muscular durante o exercício, principalmente nos segundos iniciais. PAROLIN et al. (2000) observaram significativa redução das concentrações de creatina fosfato muscular após 1 minuto de exercício a 55% do consumo máximo de oxigênio, indicando a importância deste substrato nos momentos iniciais do esforço. A glicólise anaeróbia também tem fundamental importância para a geração de ATP durante os primeiros minutos de atividade, podendo utilizar glicogênio muscular como substrato energético. Esta participação do metabolismo anaeróbio pode ser confirmada pela observação de ausência de modificações significativas do VO2 no inicio do exercício (Figura 1).
Com o prolongamento da sessão de esforço, a participação do metabolismo aeróbio torna-se mais pronunciada, não excluindo a contribuição da via anaeróbia no fornecimento energético. Contudo, em exercícios com intensidades elevadas o déficit de oxigênio pode continuar a ser observado, uma vez que o VO2 não consegue suprir a demanda.
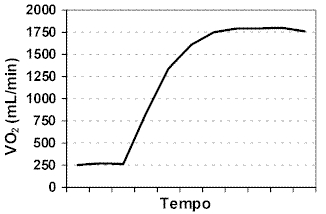
Figura 1. Consumo de oxigênio (VO2) durante o exercício físico.
Utilização de substrato energético durante o exercícioEstudos recentes têm investigado a participação das diferentes vias metabólicas para fornecimento de energia em várias condições experimentais.
RIDDELL et al. (2001) avaliaram a utilização de substrato energético durante a realização do exercício (90 minutos de exercício em cicloergômetro à 55% do VO2máx) em meninos com idade entre 10 e 14 anos, discriminando a participação das gorduras e carboidratos no fornecimento de energia. Concordando com os achados clássicos, os resultados deste experimento indicaram que a participação do metabolismo glicídico é reduzida com a passar do exercício, enquanto que a contribuição por meio da oxidação das gorduras aumenta de maneira inversa (Figura 2). Este comportamento na utilização de substrato pelas vias aeróbias e anaeróbias também foi observado após a administração de bebida à base de glicose e glicose/frutose, e refletiu em um aumento da concentração de lactato sangüíneo, principalmente nos 20 minutos iniciais do exercício onde a mobilização de glicogênio foi mais acentuada. Após o aumento inicial da lactacidemia a observação de um estado estável confirma a hipótese de “equilíbrio” metabólico promovido pela oxidação dos lipídeos.
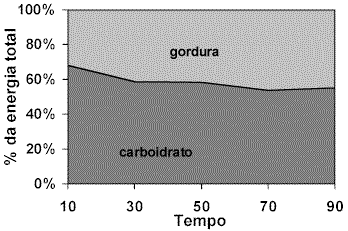
Figura 2. Porcentagem de contribuição energética proveniente de gordura e carboidrato.
Modificado de Riddell et al. (2001).A realização crônica do exercício físico, ou seja o treinamento físico, pode promover uma mudança no perfil de utilização de substratos energéticos, aumentando ou diminuindo a participação dos diferentes tipos de “combustíveis” durante a realização aguda de esforço. PUTMAN et al. (1998) investigaram a resposta de homens submetidos a 7 dias de treinamento submáximo em cicloergômetro (2h/d, 60% VO2máx) à uma sessão única de exercício. Primeiramente, na condição de repouso, observou-se que embora o período de treinamento tenha sido curto, este surtiu efeitos sobre as reservas musculares de substrato energético, promovendo aumento das reservas de glicogênio. Esta maior concentração de glicogênio no grupo treinado possivelmente deve-se aos mecanismos relacionados a supercompensação, e a adaptações que favorecem o acúmulo deste substrato como o aumento dos transportadores de glicose “GLUT4” após o treinamento (PUTMAN et al., 1998). Após a realização aguda de atividade os estoques de glicogênio permaneceram superiores no grupo treinado. Este resultado parece refletir uma mudança do perfil metabólico pela ação do treinamento, resultando em redução da glicogenólise, já que a produção de lactato sangüíneo foi menor no grupo treinado. Uma melhora da capacidade oxidativa do músculo também pode ter contribuído com a observação de concentrações mais baixas de lactato.
A utilização de glicogênio muscular durante o exercício físico pode dever-se a um aumento da secreção de hormônios contrarregulatórios os quais são responsáveis pelo catabolismo das reservas energéticas. Tem sido sugerido que aumentos da secreção de epinefrina resultam num aumento da utilização de glicogênio intramuscular devido ao aumento da atividade da glicogênio-fosforilase pela estimulação adrenérgica (RICHTER et al., 1982). FEBBRAIO et al. (1998) analisaram os efeitos da epinefrina sobre a glicogenólise muscular em homens submetidos ao exercício físico e observaram que a utilização de glicogênio é maior durante o exercício quando o grupo é submetido à infusão do hormônio. O resultado também foi acompanhado de elevações nas concentrações de lactato pós-exercício e oxidação de carboidratos, e sugerem que a utilização de glicogênio muscular durante o exercício é mediada pelos níveis circulantes de epinefrina.
A quantidade inicial de substrato energético estocado nos tecidos pode ser responsável pela antecipação ou retardo da utilização de determinadas reservas energéticas. WELTMAN et al. (1998) estudaram os efeitos dos níveis iniciais de glicogênio muscular sobre a utilização de substratos energéticos durante o exercício. Os sujeitos que apresentaram um conteúdo inicial menor de glicogênio muscular, mostraram aumento significativo das concentrações de ácidos graxos livres ao final da atividade física, indicando mobilização dos triglicerídeos estocados nos adipócitos. Novamente é destacada a relação com a secreção de epinefrina que apresentou significativo aumento durante o esforço. A participação das gorduras no fornecimento de energia do grupo com baixos níveis iniciais de glicogênio foi maior que a do grupo controle, onde a contribuição da glicose, glicogênio e lactato foi responsável pela manutenção da atividade muscular durante o exercício. JEUKENDRUP et al. (1996) observaram respostas semelhantes em ciclistas treinados submetidos ao exercício agudo (57% VO2máx.). Neste trabalho, a contribuição relativa para a geração de energia, proveniente da oxidação de gordura, foi aumentada, e a participação da oxidação de carboidratos diminuída no grupo com menores níveis iniciais de glicogênio (Figura 3).
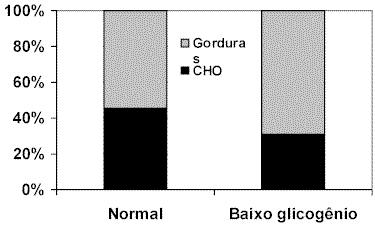
Figura 3. Fornecimento relativo de energia pela oxidação de carboidratos (CHO)
e gorduras em ciclistas com níveis normais e baixos de glicogênio muscular.
Modificado de Jeukendrup et al. (1996)PAROLIN et al. (2000) simulando uma condição de maior exigência durante o exercício, induziu homens à hipóxia e avaliou as respostas metabólicas à esta condição experimental. Manutenções das concentrações de ácidos graxos livres e glicerol foram acompanhadas de significativo aumento do lactato sangüíneo, indicando a ativação da via anaeróbia para geração rápida de ATP. A taxa de glicogenólise muscular no grupo submetido à hipóxia foi maior que a do grupo controle, tanto no início (0-1 minuto) quanto no final do exercício (15 minutos). No início do exercício, o fornecimento de ATP pela glicólise foi aumentado pela condição de hipóxia (Figura 4). Este tipo de comportamento metabólico reforça a contribuição dos componentes anaeróbios no momento inicial do exercício, onde o déficit de oxigênio limita a via aeróbia de fornecimento de energia.
Figura 4. Fornecimento de energia via creatina fosfato (CP), glicólise e fosforilação oxidativa no 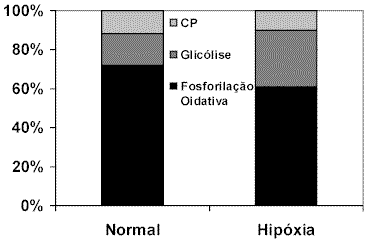
início do exercício (0-1 minuto) nas condições normal e hipóxia.
Modificado de Parolin et al. (2000)Os efeitos da intensidade do exercício sobre a utilização de substrato energético foram investigados por ROMIJN et al. (2000) em mulheres treinadas submetidas a exercício em cicloergômetro à 25, 65 e 85% do VO2máx. Os resultados indicam que a oxidação das gorduras ocorre com maior intensidade a 65% do VO2máx, embora exercícios desempenhados à 85% do VO2máx também tenham resultado em maior oxidação quando comparado com o exercício de baixa intensidade (25% VO2máx). Já a oxidação dos carboidratos ocorre em ambas intensidades (65 e 85% do VO2máx), sendo que a utilização de substrato glicídico é mais significativa à 85% do VO2máx. Os autores deixam de explorar com mais profundidade os resultados para comparar com estudos anteriores realizados com homens. Contudo, citam a participação de hormônios como epinefrina sobre a lipólise e a utilização de AGL.
Os estudos analisados nesta revisão apresentam resultados que coincidem com os achados clássicos da literatura, onde a participação da via anaeróbia de fornecimento de energia é observada principalmente nos momentos iniciais do exercício ou durante a realização de esforços com intensidades suficientemente altas que resultam em um déficit de oxigênio. Durante o “restabelecimento” do organismo frente ao exercício, a participação da via aeróbia torna-se cada vez mais significativa, utilizando componentes lipídicos em maior quantidade.
Referências bibliográficas
FEBBRAIO, M.A., LAMBERT, D.L.; STARKIE, R.L.; PROIETTO, J.; HARGREAVES, M. Effect of epinephrine on muscle glycogenolysis during exercise in trained men. J. Appl. Physiol. 84(2): 465-470, 1998.
FOX, E.L.; BOWERS, R.W.; FOSS, M.L. Fontes de energia. In: FOX, E.L.; BOWERS, R.W.; FOSS, M.L. Bases fisiológicas da Educação Física e dos Desportos. 4ed. Rio de Janeiro: Guanabara Koogan, 1991. p.10-27.
JEUKENDRUP, A.E.; BORGHOUTS, L.B.; SARIS, W.H.M.; WAGENMAKERS, A.J.M. Reduced oxidation rates of ingested glucose during prolonged exercise with low endogenous CHO availability. J. Appl. Physiol. 81(5): 1952-1957, 1996.
McARDLE, W.D.; KATCH, F.L.; KATCH, V. Transferência de energia no exercício. In: McARDLE, W.D.; KATCH, F.L.; KATCH, V. Fisiologia do exercício: energia, nutrição e desempenho humano. 3ed. Rio de Janeiro: Guanabara Koogan, 1992. p.80-93.
PAROLIN, M.L.; SPRIET, L.L.; HULTMAN, E.; HOLLIDGE-HORVAT, M.G.; JONES, N.L.; HEIGENHAUSER, G.J.F. Regulation of glycogen phosphorylase and PDH during exercise in human skeletal muscle during hypoxia. Am. J. Physiol. 278: E522-E534, 2000
PUTMAN, C. T.; JONES, N.L.; HULTMAN, E.; HOLLIDGE-HORVAT, M.G.; BONEN, A.; McCONACHIE, D.R.; HEIGENHAUSER, G.J.F. Effects of short-term submaximal training in humans on muscle metabolism in exercise. Am. J. Physiol. 275 (38): E132-E139, 1998.
RIDDELL, M. C., BAR-OR, O.; WILK, B.; PAROLIN, M.L.; HEIGENHAUSER, G.J.F. Substrate utilization during exercise with glucose and glucose plus fructose ingestion in boys ages 10-14 yr. J. Appl. Physiol. 90: 903-911, 2001
RICHTER, E. A.; RUDERMAN, N.B.; GAVRAS, H.; BELUR, E.R.; GALBO, H. Muscle glycogenolysis during exercise: dual control by epinephrine and contractions. Am. J. Physiol. 242 (5): E25-E32, 1982.
ROMIJN, J. A.; COYLE, E.F.; SIDOSSIS, L.S.; ROSENBLATT, J.; WOLFE, R.R. Substrate metabolism during different exercise intensities in endurance-trained women. J. Appl. Physiol. 88: 1707-1714, 2000.
WELTMAN, S.M.; BOSCH, A.N.; DENNIS, S.C.; NOAKES, T.D. Influence of muscle glycogen content on metabolic regulation. Am. J. Physiol. 274 (37): E72-E82, 1998.
WILMORE, J.H.; COSTILL, D.L. Basic energy systems. In: WILMORE, J.H.; COSTILL, D.L. Physiology of sport and exercise. Champaign: Human Kinetics, 1994. p.92-121.
| |
|
|---|---|
|
revista
digital · Año 8 · N° 54 | Buenos Aires, Noviembre 2002 |
|