Marcadores fisiológicos de overtraining (parte 1): análise de um protocolo
para ratos ‘wistar’ submetidos a treinamento de corrida contínua
Introdução
O processo de treinamento divide-se em períodos chamados ciclos, que podem ter
uma predominância de atividades aeróbias, que podem ser realizadas por longos
períodos de tempo (maior volume) em intensidades que variam de submáximas a
máximas (determinadas pelo VO2max ou limiar anaeróbio), características dos
treinamentos do tipo continuo ou de endurance; ou conter atividades de
menor duração, realizadas em intensidades supramáximas, com predomínio do
metabolismo anaeróbio como gerador de ATP (treinamento intermitente, de
resistência anaeróbia). Devido à alta intensidade de esforço empregada, o
treinamento do tipo intermitente requer que os exercícios sejam sempre
intercalados por pausas curtas, quando predomina o metabolismo aeróbio,
importante para facilitar a recuperação, principalmente de fosfocreatina nas
células musculares.
O início do overtraining é conhecido na literatura como overreaching
e se instala quando o repouso entre o próximo treinamento ou competição é
insuficiente (Bruin et al., 1994, Tiidus, 1998; Lehmann et al., 1998). Embora
esta condição também induza fadiga prematura, pois a recuperação é
incompleta, pode ser facilmente revertida com um ou dois dias de pouco ou nenhum
treino. Entretanto, como pouco é conhecido em relação à quantidade ótima de
treinamento e os fatores que influenciam a recuperação e supercompensação (Figura
1), ainda hoje existe um componente muito grande de empirismo na aplicação
dos métodos de treinamento, que são na sua grande maioria empregados sem muita
base científica. Ou seja, os atletas normalmente são submetidos a cargas de
esforço muito altas, com pouco tempo de recuperação, para não se pecar pela
falta de estímulo, sendo impossível prever se a carga de esforço exigida da
equipe está excedendo o limite individual de estresse que o organismo pode
suportar. Tudo isso torna o limiar entre um treinamento ideal e o overtraining
muito tênue. O maior problema é que ainda não existem descritos na literatura
marcadores completamente confiáveis, nem de adaptação, nem de overreaching,
que ajudem a diagnosticar esse limite individual de estresse que impeça a
instalação do overtraining. Além disso, a literatura é carente de
protocolos de exercícios que se preocupem em estabelecer esse quadro de
desbalanço entre exercício/repouso em animais de laboratório, o que também
dificulta o estudo da sensibilidade de biomarcadores nessa situação.
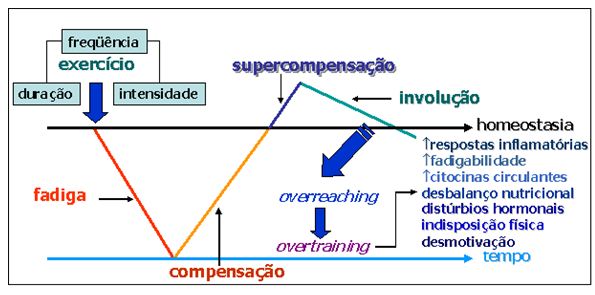
Figura
1. A influência do estímulo estressor do exercício físico nas
regulações positivas (supercompensação)
e
na instalação de estresse metabólico (overreaching) e estresse generalizado
(overtraining).
A tentativa de compreender melhor a modulação de parâmetros fisiológicos,
tais como os níveis plasmáticos de ácido úrico, creatinina e uréia, por
intermédio de experimentação animal é valida, pois há a possibilidade de
elaboração de protocolos que não seriam possíveis de controle através da
participação de seres humanos. No caso de protocolos com característica
intermitente, há poucos estudos que descrevem tal estratégia (Antunes Neto et
al., 2006; Antunes Neto et al., 2008 a,b). Assim, visamos contribuir no
aprofundamento sobre determinação de indicadores satisfatórios de
diagnóstico do quadro de overtraining.
Materiais
e métodos
Animais. Utilizamos ratos machos Wistar,
obtidos do Centro de Bioterismo da UNICAMP, com 2 meses de idade no início dos
protocolos de exercícios. Durante toda a execução dos protocolos, os ratos
foram mantidos em ambiente controlado, com temperatura de 18 - 22°C e
fotoperíodo invertido de 12 horas, estando providos de ração de laboratório
e água ad libitum.
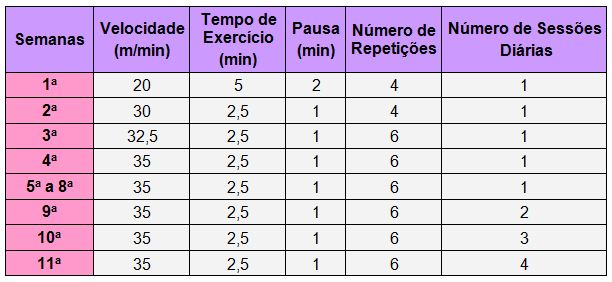
Proposta de Protocolo Intermitente de Corrida para Indução de Overtraining.
Os ratos foram submetidos a sessões de
corrida em esteira rolante automatizada com inclinação de -3°. Porém, antes
da realização do respectivo protocolo, realizamos um período de adaptação
(1 semana) do animal com o equipamento, onde eles chegaram a correr por 5
minutos numa velocidade de 10m/min. Esta semana inicial teve como finalidade
separar os animais que corriam voluntariamente daqueles que se recusavam a
correr. O protocolo de exercício intermitente (intervalado) teve onze semanas
de duração, dividido da seguinte forma:
-
primeiras
quatro semanas (adaptação): aumento na carga de esforço;
-
4a.
a 8a. semana (manutenção): exercícios diários com a mesma carga de
esforço alcançada ao final da 4a semana;
-
semana
2x/dia (9ª. Semana – incremento 1): uma semana de exercícios com a
mesma carga das semanas anteriores, 2 vezes ao dia, com intervalo de 4 horas
entre eles.
-
semana
3x/dia (10a. semana – incremento 2): uma semana com 3 sessões de
treino ao dia, com intervalo de 3 horas entre eles;
-
semana
4x/dia (11a. semana – incremento 3): uma semana com 4 sessões de
treino ao dia, com intervalo de 2 horas entre eles.
Tabela
1. Protocolo de treinamento intermitente para indução de
overtraining
-
Preparo
da Amostra. O plasma foi obtido
após centrifugação, por 10 min a 3000 rpm, do sangue restante. O plasma
foi separado e armazenado a -80°C após tratado com butilhidroxitolueno
(BHT) 90mM, este que atua como um seqüestrador específico de espécies
reativas de oxigênio.
-
Dosagens
de Parâmetros Metabólicos.
As
análises de creatinina, uréia e ácido úrico foram realizadas no aparelho
Autolab PM 4000, através de kits de análises clínicas específicos: TGO –
kit QUIMIAST, uréia e creatinina – kit QUIMIURE, ambos da Böehringer
Mannheim, e ácido úrico – kit específico Roche.
-
Apresentação
Gráfica e Análises Estatísticas.
Os
gráficos estão apresentados no modelo box plot (software S-Plus). O box
plot ordena os dados pela mediana e os divide em quartis. Ou seja, acima e
abaixo da mediana encontram-se 50% dos dados obtidos, 25% acima da mediana e
25% abaixo dela e as duas linhas tracejadas representam os 50% dos valores
mais distantes. Os dados que se diferenciam extremamente do comportamento
geral são colocados para fora dos quartis e são chamados de outliers
ou valores extremos. Optamos por apresentar os resultados obtidos na forma de box
plot por entendermos que esse tipo de gráfico descreve melhor o
comportamento do grupo de animais frente às várias situações de
exercício, permitindo uma melhor visualização da variabilidade dos dados. O
interessante desta forma de apresentação gráfica é a possibilidade de
poder visualizar a disposição de todo o grupo, conseguindo, assim, situar a
maior concentração dos resultados (Mcgill et al., 1978). As análises
estatísticas foram feitas através do programa GraphPad Instat® (San Diego,
CA). Utilizamos o teste ANOVA para amostras pareadas e, como pós-teste, foi
adotado o teste de Tukey. Valores de p<0.05 foram considerados
significativos. Vale ressaltar que a apresentação gráfica ressalta valores
da mediana, enquanto que o tratamento estatístico considera média e
desvio-padrão.
Resultados
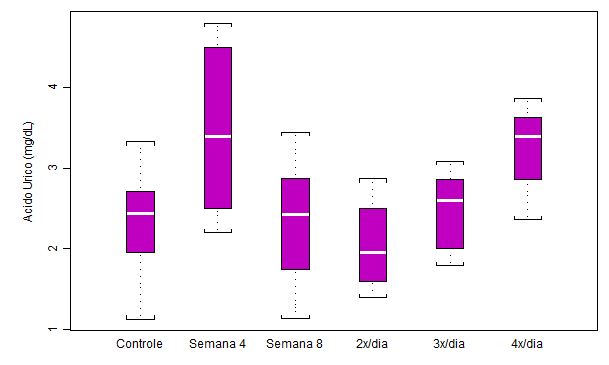
Figura
2. Concentração plasmática de ácido úrico (mg/dL) em ratos submetidos a
protocolo de treinamento
intermitente
por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada
semana)
Os dados das concentrações plasmáticas de ácido úrico (mg/dL) mostraram uma
grande variabilidade em relação a esse tipo de treinamento. O aumento na
concentração de ácido úrico na Semana 4 pode indicar que houve elevação
nos níveis de estresse oxidativo neste estágio do treinamento. As diferenças
estatísticas significativas obtidas foram: p<0.05 controle em relação a
Semana 4; p<0.05 Semana 4 em relação a Semana 8; p<0.01 Semana 4 em
relação a 2x/dia; p<0.05 2x/dia em relação a 4x/dia.
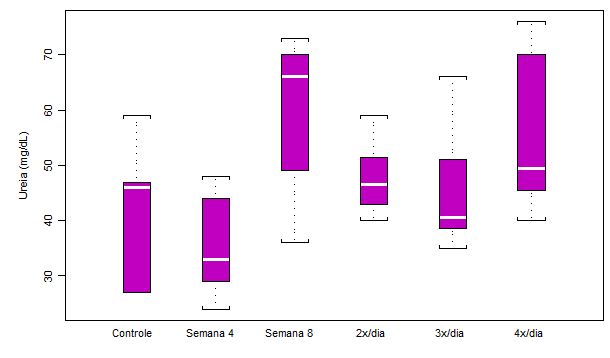
Figura
3. Concentração plasmática de uréia (mg/dL) em ratos submetidos a
protocolo de treinamento
intermitente
por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada
semana)
Os valores da mediana da concentração plasmática de uréia (mg/dL) aumentaram
a partir da análise da Semana 8, sugerindo um efeito mais tardio na taxa
catabólica protéica. O aumento mediano na Semana 8, em comparação a Semana
4, chegou a exatos 100%. Após a análise da Semana 8, a concentração de
uréia no plasma não se alterou de forma tão acentuada, mesmo com o aumento na
freqüência diária dos exercícios de corrida. Os valores obtidos de
significância estatística foram: p<0.01 Semana 4 em relação a Semana 8;
p<0.05 Semana 4 em relação a 4x/dia.
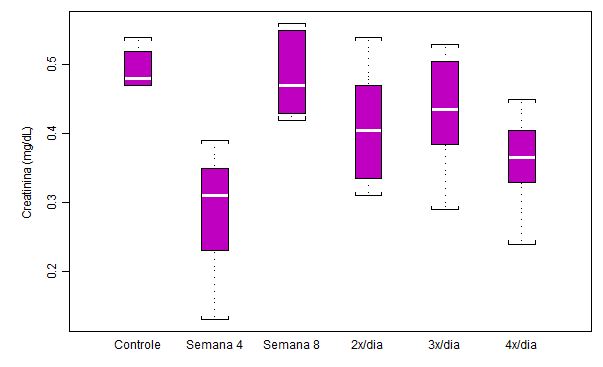
Figura
4. Concentração plasmática de creatinina (mg/dL) em ratos submetidos a
protocolo de treinamento
intermitente
por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada
semana)
Da mesma forma que o observado para o treinamento contínuo (parte 1), a
concentração de creatinina (mg/dL) diminuiu drasticamente somente após as
primeiras quatro semanas (Semana 4) de exercício intermitente, voltando a
valores próximos do grupo controle na oitava semana (Semana 8). Tal
diminuição na Semana 4, em relação a mediana do grupo controle, chegou a
35.5.%. Na fase onde houve a tentativa de indução de overtraining
(análises 2x/dia, 3x/dia e 4x/dia), obtivemos uma grande dispersão dos
resultados, tal como ocorrido na Semana 4, porém com os valores de mediana
flutuando numa mesma faixa. Esses dados sugerem que o estresse dessas últimas
três semanas de treinamento também não foi predominantemente metabólico. Os
valores de significância estatística foram: p>0.001 controle em relação a
Semana 4; p<0.001 Semana 4 em relação a Semana 8; p<0.05 Semana 4 em
relação a 2x/dia; p<0.01 Semana 4 em relação a 3x/dia; p<0.05 controle
em relação a 4x/dia; p<0.05 Semana 8 em relação a 4x/dia.
Discussão
Tal como no treinamento contínuo (artigo Parte 1), utilizamos as análises de
concentração plasmática de ácido úrico como um indicador da condição
antioxidante plasmática. Vimos que a hipoxantina é convertida a ácido úrico
via ativação da enzima xantina oxidase durante exercícios de alta intensidade
(Hellsten et al., 1988), condição que caracteriza o nosso protocolo de
treinamento intermitente (Figura 5).
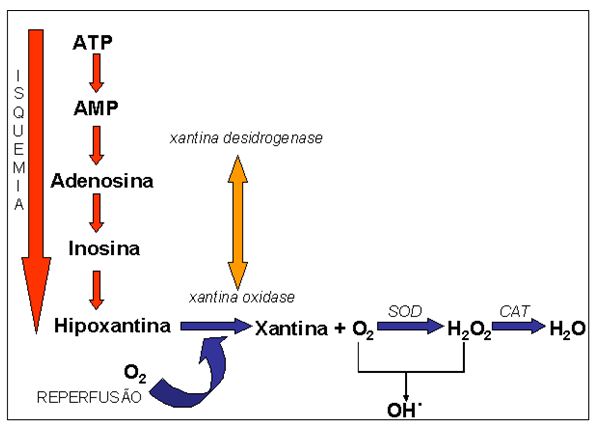
Figura
5. Mecanismo de produção de espécies reativas de oxigênio através do
processo isquemia-reperfusão. Adaptado de Adkison et al. (1986) e Granger et
al. (1986)
Os resultados mostraram que o treinamento intermitente induziu aumento
considerável na concentração plasmática de ácido úrico no momento de
implementação da maior sobrecarga de corrida (análises da Semana 4 e 4x/dia).
Considerando as características metabólicas dos dois tipos de protocolos, o
treinamento intermitente possui uma tendência maior de gerar um quadro de
isquemia-reperfusão, pelo fato dos exercícios de corrida de alta intensidade
(condição de anóxia) serem intercalados com períodos recuperativos de pausa
passiva (condição de oxigenação). A literatura aponta que a ativação da
enzima xantina oxidase ocorre principalmente em eventos onde há
isquemia-reperfusão da célula (Adkison et al., 1986; Granger et al., 1986).
Os marcadores de alterações metabólicas - uréia e creatinina – tiveram um
comportamento muito próximo ao obtido pelo treinamento contínuo (artigo Parte
1), mostrando grande dispersão dos resultados. Bruin e colaboradores (1994),
apesar de terem observado várias alterações metabólicas em cavalos de
corrida submetidos a exaustivos treinamentos de endurance e intervalado,
que os permitiram classificá-las como overtraining, não obtiveram
modificações significativas na concentração plasmática de uréia, por
exemplo. Não encontramos trabalhos na literatura relacionando alterações de
concentração plasmática de creatinina e protocolos de treinamento
intermitente. Por isso que se faz importante a utilização de vários
marcadores de estresse, sejam de ordem fisiológica ou bioquímica, pois o
heterocronismo das respostas orgânicas aos agentes estressores torna-se um
fator a mais de dificuldade de detecção da síndrome do supertreinamento.
Utilizando-se dos mesmos protocolos para estudos do comportamento de geração
de espécies reativas de oxigênio (EROs), pudemos observar que marcadores como
as enzimas antioxidantes (catalase e glutationa redutase) e produtos de
oxidações celulares de lipídios (substâncias reativas ao ácido
tiobarbitúrico – TBARS) e proteínas (proteínas carboniladas) são
extremamente mais sensíveis a modulações decorrentes dos estímulos
específicos dos procedimentos de treinamento (Antunes Neto et al., 2008 a,b).
As EROs, também conhecidas como radicais livres, são formadas, principalmente,
pela redução incompleta do oxigênio nos processos de geração de energia (Figura
6). O oxigênio consumido tem como principal via de metabolismo o sistema
aeróbio, ou seja, a mitocôndria (Antunes Neto et al., 2006). Esse sistema é
responsável pela utilização de 85 a 90% de todo o oxigênio consumido, com os
outros 10 a 15% sendo utilizados por enzimas oxidases e oxigenases e por
reações químicas de oxidação direta. Dos 85 a 90% de oxigênio que chega a
cadeia de transporte de elétrons, 2 a 5% são reduzidos unilateralmente em
metabólitos, tal como as EROs (Schneider, Oliveira, 2004). Durante a atividade
física o consumo de oxigênio pode aumentar em até 20 vezes e a captação do
mesmo pelos músculos ativos pode aumentar em até 100 vezes, favorecendo, dessa
maneira a formação de EROs (Antunes Neto et al., 2005).
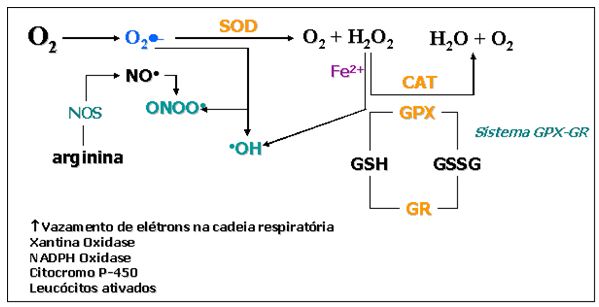
Figura
6. Mecanismos de formação de espécies reativas de oxigênio e as
estratégias enzimáticas antioxidantes. Nota-se que o aumento de consumo de
oxigênio possibilita vazamento
de
elétrons na cadeia respiratória mitocondrial, o que potencializa a geração
das espécies reativas. Enzimas como xantina oxidase, NADPH oxidase, bem como os
eventos
decorrentes
de processos inflamatórios (ativação leucocitária), contribuem para
geração elevada dos radicais livres. Uma das vias musculares mais
potentes
de geração de peróxinitrito é a degradação do aminoácido arginina,
circunstância vista em condição de exercício exaustivo
O estresse oxidativo ocorre em circunstancias nas quais há desequilíbrio entre
os sistemas prooxidantes e antioxidantes, de maneira que os primeiros sejam
predominantes (Scheneider, Oliveira, 2004). Dentro de uma estratégia de
manutenção do estado redox contra condições oxidantes, o sangue exerce um
papel fundamental, fazendo o transporte e redistribuição dos antioxidantes
para todo o corpo; dessa maneira, a capacidade antioxidante no sangue pode nos
dar estimativas dos níveis de estresse oxidativo, permitindo um modo de
mensuração menos invasivo que por outras vias, como pela biopsia (Antunes Neto
et al., 2006). Para livrar o organismo dos efeitos deletérios das EROs, há
sistemas antioxidantes divididos em duas classes: o sistema não enzimático,
composto principalmente por β-caroteno (pro vitamina A), ácido ascórbico
(vitamina C), α-tocoferol (vitamina E) e grupamentos sulfidrila (SH), e o
sistema enzimático, com a catalase (CAT), glutationa peroxidase (GPX),
glutationa redutase (GR) e superóxido desmutase (SOD) como as principais
enzimas (Powers et al., 1994).
Do ponto de vista metodológico, podemos pontuar a dificuldade em se obter
adaptações positivas através de um protocolo intermitente de corrida
utilizando-se animais. Um fator chave ao longo do processo de treinamento é
manter efetivamente os animais correndo na esteira. Como o tempo de corrida de
cada exercício é curto, qualquer descuido do animal pode interferir no
resultado das análises. Muitas vezes, o animal consegue permanecer sem correr
por ficar encostado na parte posterior da esteira, mesmo esta estando em
rotação. Essa posição pode causar lesões em articulações e músculos,
sendo um outro fator que pode contribuir com interpretações indevidas dos
resultados.
Concluindo, tal como relatado para o protocolo de treinamento de corrida
contínua, evidenciamos que o protocolo intermitente não possibilita uma clara
tendência de modificações fisiológicas (ácido úrico, creatinina, uréia),
o que nos leva a ressaltar a importância de eventos moleculares, sobretudo a
geração de EROs e suas reações conseqüentes, para a instalação de um
quadro de overtraining.
Referências
bibliográficas
-
ADKISON,
D.; HÖLLWARTH, M. E.; BENOIT, J. N. Role of free radicals in
ischemia-reperfusion injury to the liver. Acta Physiologica Scandinavica,
v. 126, suppl. 548, p. 101-107, 1986.
-
ANTUNES
NETO, J. M. F.; TOYAMA, M. H.; CARNEIRO, E. M.; BOSCHERO, A. C.;
PEREIRA-DA-SILVA, L.; MACEDO, D. V. Circulating leukocyte heat shock protein
70(hsp70) and oxidative stress markers in rats after a bout of exhaustive
exercise. Stress, v. 9, n. 2, p. 107-115, 2006.
-
ANTUNES
NETO, J. M. F.; PEREIRA-DA-SILVA, L.; MACEDO, D. V. Heat shock proteins as
oxidative stress markers in rats submitted to exhaustive intermittent
running training. Brazilian Journal of Biomotricity, v. 2, n. 3, p.
160-175, 2008.
-
ANTUNES
NETO, J. M. F.; PEREIRA-DA-SILVA, L.; MACEDO, D. V. Proteínas de estresse
“HSP70” atuam como marcadoras de estresse oxidativo em ratos “Wistar”
submetidos a treinamento contínuo de corrida para indução de “overreaching”.
Revista Brasileira de Educação Física, Esporte, Lazer e Dança, v.
3, n. 4, p. 116-130, 2008.
-
ANTUNES
NETO, J. M.F.; PEREIRA-DA-SILVA L.; MACEDO, D. V. Biomarcadores de estresse
oxidativo: novas possibilidades de monitoramento em treinamento físico. Revista
Brasileira de Ciência e Movimento, v. 13, n. 2, p. 73-79, 2005.
-
BRUIN,
G.; KUIPERS, H.; KEIZER, H. A.; VANDERVUSSE, G. J. Adaptation and
overtraining in horses subjected to increasing training loads. Journal of
Applied Physiology, v. 76, n. 05, p. 1908-1913, 1994.
-
GRANGER,
D. N.; HÖLLWARTH, M. E.; PARKS, D. A. Ischemia-reperfusion injury: role of
oxygen-derived free radicals. Acta Physiologica Scandinavica, v. 126,
suppl. 548, 47-63, 1986.
-
HELLSTEN,
Y.; AHLBORG, G.; JENSEN-URSTAD, M. Indication of in vivo xanthine oxidase
activity in human skeletal muscle during exercise in man. Acta
Physiologica Scandinavica, v. 137, p. 341-345, 1988.
-
LEHMANN,
M.; FOSTER, C.; DICKHUTH, H. H.; GASTMANN, U. Autonomic imbalance hypothesis
and overtraining syndrome. Medicine and Science in Sports and Exercise,
v. 30, n. 07, p. 1140-1148, 1998.
-
MCGILL,
R.; TUKEY, J. W.; LARSEN, W. A. Variations of Box Plots. American
Statistician, v. 32, n. 01, p. 12-16, 1978.
-
POWER,
S. K.; CRISWELL, D.; LAWLER, J.; JII, LL.; MARTIM, D.; HERB, R. A.; et al.
Influence of exercise and fiver type on antioxidante enzyme activity in rat
skeletal muscle. American Journal of Physiological, v. 266, p.
R375-R380, 1994.
-
SCHNEIDER,
C. D.; OLIVEIRA, A. R. Radicais livres de oxigênio e exercício: mecanismos
de formação e adaptação ao treinamento físico. Revista Brasileira de
Medicina do Esporte, v. 10, n. 4, p. 308-313, 2004.
-
TIIDUS,
P. M. Radical species in inflamation and overtraining. Canadian Journal
of Physiology and Pharmacology, v. 76, p. 533-538, 1998.
Outros artigos em
Portugués
 |
|
|
EFDeportes.com, Revista Digital · Año 17 · N° 171 | Buenos Aires,
Agosto de 2012
© 1997-2012 Derechos reservados
|
