
|
|||
|---|---|---|---|
|
|
Os estímulos moleculares geradores para produção de força muscular Los estímulos moleculares generadores para la producción de la fuerza muscular The molecular stimulus generators to produce muscle strength |
|
|
|
*Doutor em Biologia Funcional e Molecular, pelo Departamento de Bioquímica Instituto de Biologia, Universidade Estadual de Campinas, UNICAMP **Professor da Faculdade de Educação Física, Departamento de Estudos da Atividade Física Adaptada, Universidade Estadual de Campinas, UNICAMP |
Joaquim Maria Ferreira Antunes Neto* Roberto Vilarta** (Brasil) |
|
|
|
Resumo A literatura aponta em grande volume dados sobre quais estímulos atuam nos eventos ligados à síntese protéica. A maioria dos estudos está voltada para a área da biologia molecular e sinalizadores nucleares. O objetivo principal deste artigo é mostrar o início das fases de adaptação ao treinamento de força, sobretudo aquelas relacionadas à tensão mecânica, que pode gerar um distúrbio nas estruturas sarcoméricas e, por fim, moleculares. Unitermos: Força muscular. Treinamento de força. Sarcômero. Adaptação. Sinalização molecular.
Abstract The literature points to a large volume of data on which literary stimuli act on events related to protein synthesis. Most studies are focused on the area of molecular biology and nuclear flags. The aim of this paper is to show the beginning stages of adaptation to strength training, especially those related to mechanical stress, which can generate a disturbance in the sarcomeric structures and, finally, molecular events. Keywords: Muscle strength. Strength training. Sarcomere. Adaptation. Molecular signaling.
|
|||
|
|
EFDeportes.com, Revista Digital. Buenos Aires, Año 16, Nº 164, Enero de 2012. http://www.efdeportes.com/ |
|
|
1 / 1
Introdução
Um dos mecanismos adaptativos que mais chama o interesse dentro das bases biológicas da atividade física é o que diz respeito ao processo indutor de aumento em força muscular. Smith e Rutherford (1994), em experimento que comparava as formas de adaptação ao treinamento excêntrico e concêntrico de resistência de força muscular, com duração de 20 semanas e unidades de treino por 3 vezes semanais, submeteram seus voluntários a um treinamento de 4 séries de 10 repetições com uma carga de peso que variava de 60% - 80% da repetição máxima, onde cada perna executava uma forma de contração muscular durante o período de treinamento. Os resultados mostraram que não houve diferença significativa no aumento de área de secção transversal entre as duas pernas; contudo, na perna treinada concentricamente, ocorreu um aumento significativo em força isométrica. Desta forma, o elevado custo metabólico das contrações concêntricas poderia ter desempenhado um importante papel enquanto estímulo para o aumento em força muscular. Porém, vale ressaltar que a forma de teste empregada neste trabalho não condiz com o método de treinamento utilizado pelos grupos concêntrico e excêntrico, o que contraria a teoria de especificidade de treinamento.
Nesta mesma tendência, Jones e Rutherford (1987), estudando os efeitos de três regimes diferentes de treinamento de força e a natureza das mudanças resultantes, em um período de 12 semanas com unidades de treino 3 vezes semanais de 4 séries de 6 repetições, realizaram o seguinte protocolo: o grupo isométrico mobilizou uma carga de peso de 80% de 1 repetição máxima (1 RM) unilateralmente, com cada contração isométrica durando 4s e tendo 2s de intervalo entre as contrações; o treinamento excêntrico e concêntrico foi realizado por cada voluntário selecionado para estes 2 tipos de contrações, onde cada perna executou uma modalidade de contração, com carga de 80% de 1 RM concêntrica para o grupo concêntrico e carga de 145% de 1 RM concêntrica para o grupo excêntrico. O treinamento isométrico resultou em um aumento na força isométrica de 35 ± 19%, enquanto que, para o treinamento excêntrico e concêntrico, os aumentos em força isométrica foram de 11 ± 36% e 15 ± 80%, respectivamente. Estes valores indicam que nenhuma modalidade de treinamento - excêntrica e concêntrica - é mais efetiva do que a outra, mesmo apesar do treinamento excêntrico ter sido realizado com uma carga aproximadamente 50% mais elevada em comparação ao treinamento concêntrico. Por outro lado, contrações concêntricas possuem um maior custo energético - fator aparentemente decisivo para aumento em força muscular, segundo Smith e Rutherford (1994) - e, mesmo assim, obteve-se ganhos em força similares ao treinamento excêntrico. Se, desta forma, o estímulo para um aumento em força fosse o elevado estresse mecânico ou o custo metabólico, uma ou outra forma de treinamento poderia ter sido mais efetiva. Mas isso não foi o caso, sendo sugerido que nenhum desses fatores, isoladamente, seja único para o desencadeamento das adaptações musculares. O estímulo pode estar relacionado a uma combinação de fatores mecânicos e metabólicos ou, então, a um limiar de estresse mecânico (Jones, Rutherford, 1987).
Apesar do estímulo crítico para aumento em força muscular ainda não esteja totalmente definido, existe um consenso favorável em considerar fator essencial um determinado nível de estresse mecânico (Goldberg et al., 1975). Postula-se que pode ser a quantia de tensão muscular desenvolvida a chave da resposta que, de certo modo, origina o sinal para o aumento rápido de aminoácidos e a elevação da síntese de proteínas contráteis (Colliander, Tesch, 1987). Neste caso, contrações excêntricas levam vantagem através deste mecanismo de aumento em força muscular, pois quando o “input” neural é o mesmo para ações excêntricas e concêntricas, a força registrada é muito maior na condição excêntrica. A “força adicional” gerada durante ações excêntricas parece, portanto, ser gerada por outro mecanismo que não seja determinado por recrutamento de unidades motoras ou modulação de freqüência (Tesch et al., 1990). A contribuição dos componentes elásticos do tecido muscular tem sido enfocada neste processo de geração adicional de força em condição excêntrica, em conjunto com as características de rompimento de ligações eletrostáticas entre filamentos de actina e miosina (Bennett, Stauber, 1986).
De acordo com a teoria da ponte cruzada, a força ativa gerada por uma única ponte cruzada no sarcômero resulta da ligação entre actina e miosina pela retenção eletrostática de alta afinidade, seguida pelo movimento de ponte cruzada (tensão e encurtamento) e da dissociação de actina e miosina na presença de ATP (relaxamento). Quando o músculo é ativado pelos processos do acoplamento excitação-contração, há a retirada da inibição imposta pela tropomiosina, processo esse mediado pela interação de cálcio com troponina. Formam-se, daí, os eventos mecânicos de ação entre as pontes cruzadas dos sarcômeros. A Figura 1 mostra possíveis vias de geração e transmissão de força no músculo esquelético.
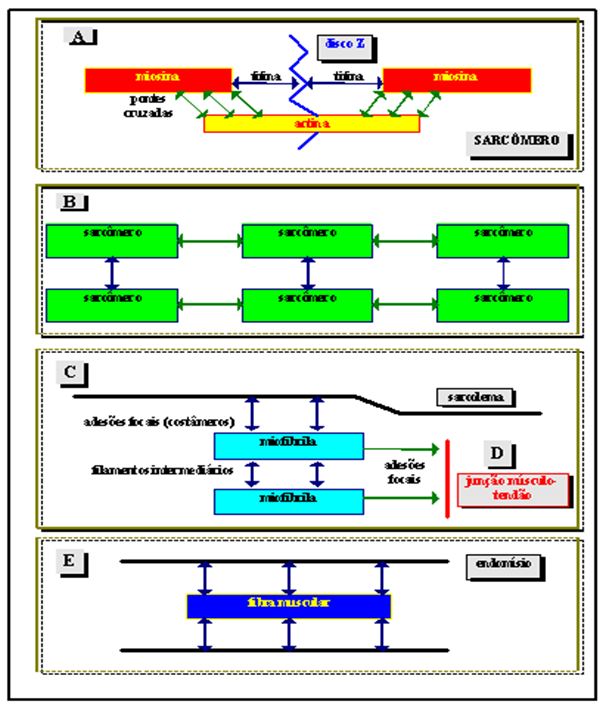
Figura 1. Vias potenciais de geração e transmissão de força no músculo. Adaptado de Patel e Lieber (1997). As flechas verdes
significam as vias de transmissão de força mais aceitas, enquanto que as flechas azuis constituem-se nas áreas recentes de pesquisa
Observe na Figura 1 que a geração de força inicia-se entre os filamentos de actina e miosina (quadro A), sendo transmitida para os discos Z e as moléculas de titina. Após, a força é transmitida em série entre os sarcômeros e, supostamente, lateralmente pelo sistema de filamentos intermediários (quadro B). O quadro C mostra a seqüência seguinte de transmissão da força, que se dá lateralmente entre as miofibrilas pelos filamentos intermediários e para a superfície da membrana pelos costâmeros (canecões entre as proteínas intracelulares e o espaço extracelular, onde a proteína vinculina é a mais presente) e pelo complexo distrofina (importante na manutenção da integridade da fibra muscular, possivelmente possuindo um comportamento estabilizador de membrana contra ruptura e degeneração). A força é transmitida para a junção miotendinosa (quadro D) por meio de canecões de adesão especializadas e para a porção externa do tendão (quadro E) através da força escoada lateralmente entre as fibras musculares em direção à matriz do tecido conjuntivo (Patel, Lieber, 1997). No caso de contrações excêntricas, pode-se postular que haja uma alteração no padrão de transmissão de força através das estruturas do sistema músculo-esquelético, o que causaria, talvez, um maior estresse mecânico principalmente aos tecidos conjuntivos e estabilizadores, estes que ficariam numa condição excedente elevada.
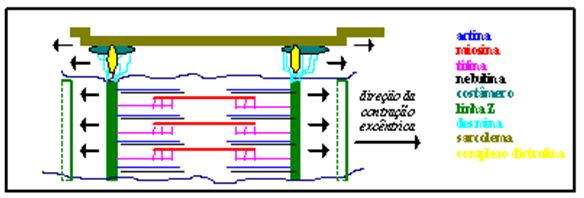
Figura 2. Padrão de transmissão de força muscular, como possível indutor de estímulo para síntese proteica
A relação existente entre contração excêntrica e produção de força, através da análise da teoria da ponte cruzada, prediz que, se imediatamente após a reação de ligação a ponte cruzada é forçadamente puxada para trás, a junção actomiosina poderá se romper antes da transdução de energia ocorrer (Figura 2). Esta separação da ponte cruzada tende a requerer mais força do que a registrada no ciclo normal contrátil (Stauber, 1989).
Processo de Síntese Protéica
De acordo com Williams (1989), muitas das respostas adaptativas importantes podem ser consideradas como questões de regulação gênica (Figura 3). A intensidade do funcionamento das estruturas celulares pode, assim, determinar o grau de atividade do aparelho gênico:
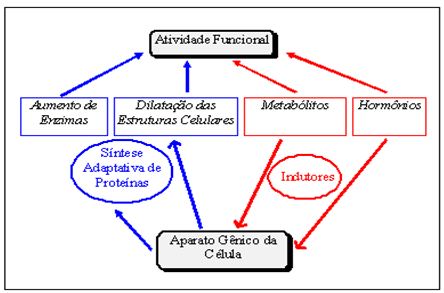
Figura 3. Mecanismos adaptativos de síntese protéica induzidos pela capacidade funcional celular. Adaptado de Viru (1994)
Um estímulo específico tende a induzir a síntese de proteínas. Pressupõe-se que o treinamento físico causa um acúmulo de metabólitos os quais, especificamente, induzem a síntese adaptativa de estruturas e enzimas relacionadas às exigências mais ativas em termos estruturais e metabólicos da célula. Em adição, mudanças hormonais são induzidas pelo treinamento, ampliando os efeitos indutores dos metabólitos e assegurando o suprimento de síntese protéica. Como resultado, uma efetiva renovação e espessamento de estruturas protéicas e um aumento em número de moléculas enzimáticas ocorrem.
No caso do aumento em síntese de proteínas do metabolismo oxidativo, pressupõe-se que contrações repetitivas do músculo-esquelético resultam na geração de um “sinal molecular (X)”, e que o acúmulo deste sinal propicia a elevação de síntese protéica específica (Williams, 1989). Algumas hipóteses são lançadas para explicar a teoria do sinal molecular:
-
O sinal X atua localizadamente.
O sinal X persiste após a atividade contrátil.
Os efeitos do sinal X são limitados às proteínas do metabolismo oxidativo e não afetam proporções globais de síntese de proteína.
As respostas adaptativas para a ocorrência de hipertrofia muscular não são explicadas pela Teoria do Sinal X, pois, diferentemente da síntese de proteínas oxidativas, parece haver uma influência de fatores tróficos para o aumento de massa muscular. O estímulo primário para a elevação de aminoácidos, de acordo com a literatura, relaciona-se com a quantia de tensão desenvolvida pelo músculo, conforme já relatado anteriormente.
No caso do exercício excêntrico, ao mesmo tempo em que sua característica lesiva assegura o início de uma resposta inflamatória, talvez por intermédio de proteases ativadas por cálcio (Croall, Demartino, 1991), o processo regenerativo parece ocorrer em condição simultânea, como se inflamação/regeneração fossem parte de um mesmo complexo de reparo, haja visto a atividade de células satélites, células fagocíticas e células linfoides nos mecanismos de manutenção e regulação das funções celulares (Stauber et al., 1988). Portanto, a extensão da ruptura homeostática nas fibras musculares submetidas ao exercício excêntrico, por intermédio de proteoglicanas, pode sinalizar a proliferação de novas miofibrilas, a regulação do processo miogênico, a fusão entre células satélites e mioblastos e a orientação de miotubos (Fritz, Stauber, 1988).
Quadro 1. Resumo dos eventos adaptativos durante o processo de regeneração tecidual. Adaptado de Hurme e Kalimo (1992)

Wong e Booth (1990a) estudaram o metabolismo de proteínas em ratos em decorrência de estimulação elétrica e atividade contrátil concêntrica (estudou-se o músculo gastrocnêmio) e excêntrica (estudou-se o músculo tibial anterior). Os protocolos foram os mesmos, dotados de várias combinações de exercícios, com estímulos de baixa, moderada e alta intensidades. O que poderia explicar o maior grau de hipertrofia, em decorrência do treinamento excêntrico, seria um aumento agudo mais prolongado em síntese protéica no músculo tibial anterior em relação ao músculo gastrocnêmio, conclusão esta obtida pela análise das medidas de RNA e proteínas específicas de RNA mensageiro. Estas diferenças em proporção de síntese protéica e massa muscular podem ser, provavelmente, devido à maior tensão específica imposta ao músculo tibial anterior durante os exercícios e treinamento (Wong, Booth, 1988), ou pela ativação mais acentuada de mecanismos degradativos sob condição concêntrica. Vale ressaltar, ainda, o trabalho de Chesley e colaboradores (1992), fazendo uma análise do aumento em síntese protéica muscular em humanos, o qual relaciona as variáveis de intensidade e volume de exercício, o músculo ou grupos musculares envolvidos, o tipo de contração muscular realizada e o estado de treinamento do indivíduo como fatores importantes em condição de gerar estímulo para aumento em síntese de proteínas na execução de um único turno de exercícios.
A análise do trabalho de Wong e Booth (1990b) permite concluir que múltiplos sítios de controle (pré-traducional, traducional e pós-traducional) podem ser estimulados pelo treinamento (Figura 4), haja visto que os aumentos após o programa de 10 semanas de treinamento foram de 41% em a-actina mRNA, de 38% no total de RNA e de 28% em proteínas no músculo tibial anterior.
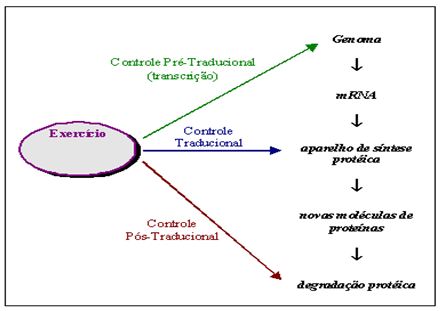
Figura 4. Múltiplos sítios de controle do processo adaptativo de síntese protéica. Adaptado de Viru (1994)
Millward (1980) aponta que o músculo grande dorsal (porção anterior) alongado por um peso adicional, em modelo animal, hipertrofia-se e desenvolve tensão via reflexo de alongamento. Pelo fato do alongamento induzir aumento em massa muscular mesmo na ausência de algum suporte hormonal ou nutricional, ele pode ser visto como o mais importante fator conhecido em regulação de crescimento muscular. Como uma condição de estresse mecânico pode levar a uma resposta bioquímica, tal como o aumento de síntese protéica, ainda não está totalmente elucidada. Uma possível ligação bioquímica entre alongamento e resposta anabólica pode ser o aumento em transporte de aminoácidos como resultado de alguma alteração na membrana celular e de seus receptores. A ocorrência de um processo de degradação protéica e acúmulo de colágeno no tecido muscular neste modelo de hipertrofia induzida pelo alongamento, juntamente com o aumento no processo de síntese desencadeado por estimulação de células satélites e mitose, parece relacionar-se com células em estado de crescimento (esquema 24). O que se postulou foi que o estímulo mecânico do alongamento desencadeou uma “cascata sinalizadora de eventos” originada de receptores ou canais localizados sobre o sarcoplasma das fibras musculares e que conduziu às modificações pós-traducionais de “fatores de resposta presentes no plasma”, propiciando, com isso, um aumento em interação proteína-proteína e alteração no processo de regulação genética da célula (Carson, 1977).
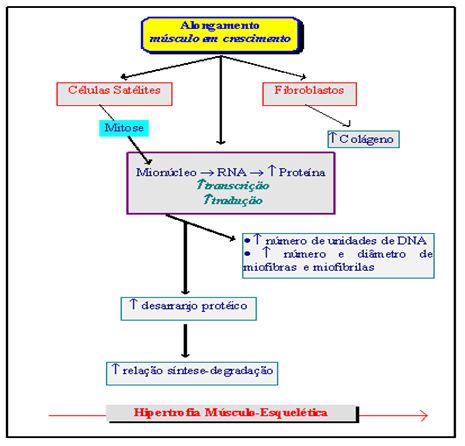
Figura 5. Possíveis eventos durante hipertrofia musculoesquelética induzida por alongamento. Adaptado de Millward (1980)
Como se pode perceber na Figura 5, a hipertrofia musculoesquelética é um evento complexo, sendo proposto um modelo para regulação na expressão genética (Carson, 1997). O modelo é composto de dois estágios de adaptação, onde os fatores determinantes para o desencadeamento de respostas hipertróficas relacionam-se com as condições de sensibilidade do músculo à sobrecarga (limiar de ativação) e com as condições micromoleculares que vêm a possibilitar ao músculo um grau de enlargamento e alteração na forma celular (a alteração no formato da célula pode ser um sinal ao núcleo para desencadear mudanças na expressão genética).
No primeiro estágio de hipertrofia, logo no início de sensibilização à sobrecarga, há um aumento na atividade do RNA. As evidências experimentais sugerem que o aumento em síntese protéica no início de hipertrofia seja um forte reflexo deste aumento de atividade do RNA, o que é possibilitada pela melhoria em eficiência na capacidade de tradução protéica (o aumento em quantidade de RNA ribossômico determina a realização desta análise). No segundo estágio de hipertrofia, nota-se um aumento na quantidade de mRNA, refletindo melhoria na capacidade de transcrição proteica e/ou aumento em estabilidade do mRNA. Esta melhoria em condição transcricional dos músculos pode ser aumentada pela adição de núcleos derivados de células satélites que se fundem com as células em hipertrofia. A capacidade de transcrição é aumentada quando os mecanismos traducionais e pós-traducionais não podem sustentar a proporção de síntese protéica requerida para o enlargamento da fibra muscular. Em adição à regulação traducional e pós-traducional, o músculo hipertrofiado poderá ter um aumento na capacidade de transcrição por mionúcleo.
Neste momento, torna-se importante retornar à discussão entre relação entre as respostas neurais e hipertróficas em virtude do treinamento de força. As alterações no processo de síntese protéica, tais como melhoria em eficiência dos eventos traducionais e transcricionais, parecem surgir com mais preponderância num segundo momento da ação periodizada de treinamento. No início do processo de treinamento de força, a resposta mais acentuada observada é um aumento da atividade eletromiográfica dos grupos musculares envolvidos com os exercícios, o que reflete adaptação neural. Os possíveis mecanismos de adaptação neural podem incluir um aumento na ativação dos músculos motores primários em um movimento específico, bem como mudanças apropriadas na ativação dos músculos sinergistas e antagonistas (Sale, 1988).
Qual seria a causa do possível aumento em ativação neural dos músculos durante treinamento de força? Komi (1986) coloca que muitos fatores podem contribuir: aumento no número de unidades motoras recrutadas, aumento na freqüência de ativação das unidades motoras e aumento na sincronização de ativação das unidades motoras. Em relação ao aumento de sincronização das unidades motoras, há duas possibilidades para explicar: os dendritos dos motoneurônios a podem receber aumento na entrada (“inputs”) das fibras sensoriais e/ou os centros motores altos aumentam a atividade das vias descendentes. Komi (1986) também sugere que o aumento na ativação dos motoneurônios-a com o concomitante aumento na sincronização das unidades motoras não somente causam melhoria nos níveis de força no primeiro momento de treinamento, mas podem também servir como um importante estímulo para os fatores hipertróficos esperados com a continuidade do processo de treinamento (Figura 6).
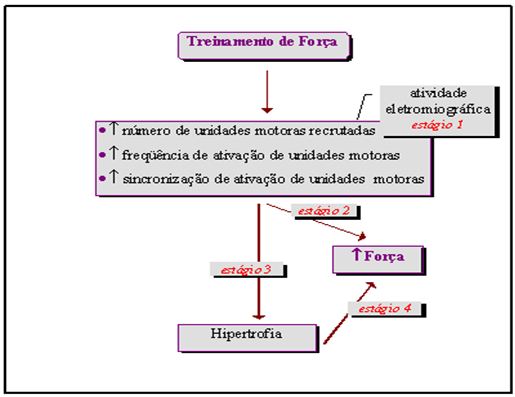
Figura 6. Representação da seqüência dos eventos que propiciam aumento de força muscular. Adaptado de Komi (1986)
A questão das respostas hipertróficas procedentes de atividades onde há alongamento ativo dos sarcômeros parece ser um processo dos mais complexos. Analisando a literatura, observam-se pressupostos de interação entre mediadores neurotróficos e intracelulares. O neurotrofismo refere-se à influência sustentada que um elemento biológico (neurônio) exerce diretamente sobre outro (fibras musculares). A estrutura interna de um neurônio é bem complexa, suportando um rápido sistema de transporte para movimentação de materiais. O transporte de materiais pode ocorrer do soma à placa motora final (transporte ortógrado) e na direção reversa (transporte retrógrado).
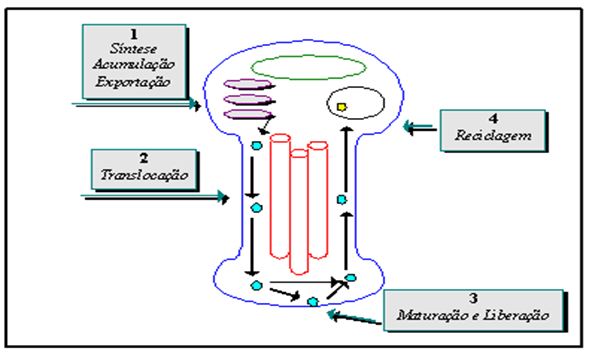
Figura 7. Sistema de transporte axonal. Adaptado de Enoka (1994)
O material transportado inclui elementos estruturais, proteínas, RNA e aminoácidos. Por exemplo, um importante passo na transferência eletroquímica de informação do neurônio ao músculo é a liberação de neurotransmissores pela vesícula em direção à fenda sináptica. Após um certo número de ciclos, a membrana da vesícula requererá reparo e tal ocorrerá no soma; a vesícula será subseqüentemente enviada de volta para a junção neuromuscular via transporte ortógrado (Enoka, 1994). No caso do processo de indução de síntese de proteínas miofibrilares, algumas neurotrofinas podem ganhar acesso ao interior da fibra muscular através de endócitos. Motoneurônios que não são ativados freqüentemente (unidades tipo IIB) podem gerar neurotrofinas que se unem a endócitos ou receptores de ligação; tal união pode também possuir afinidade com um sistema de segundos mensageiros que ativará propriedades gênicas no interior da célula. Já no caso de motoneurônios ativados freqüentemente (unidades tipo I, IIA), o impulso de atividade parece ser um fator decisivo, em conjunto com a atividade de alongamento dos sarcômeros, na indução de síntese protéica miofibrilar. Os fatores solúveis liberados pelo alongamento podem atuar sobre o núcleo de forma direta, como no caso das prostaglandinas, ou então indiretamente através de segundos mensageiros, conforme ilustra a Figura 8. O cálcio poderá ser utilizado também como um segundo mensageiro (McComas, 1996).
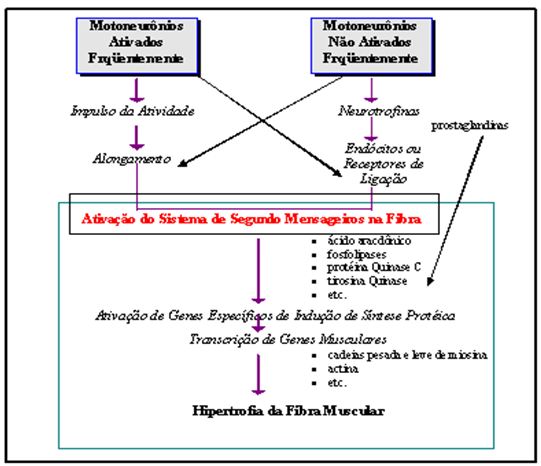
Figura 8. Mecanismo de indução de síntese protéica miofibrilar através de fatores neurotróficos e intracelulares. Adaptado de McComas (1996)
Existem genes que são ativados primeiramente para o desenvolvimento do evento de síntese protéica, tal como o gene c-fos. Estes genes, em parte, controlam a transcrição de outros genes no núcleo, incluindo aqueles que codificam para todas as proteínas especiais necessárias para a transformação das fibras musculares (cadeias pesada e leve de miosina, troponina e outras proteínas de ligação ao cálcio e várias enzimas musculares). Assim, em um estágio final, poderá haver hipertrofia da fibra muscular (McComas, 1996).
Atividade muscular excêntrica e postura
Kirby e colaboradores (1992) mostram o potencial do exercício excêntrico como um efetivo método para auxiliar em situação de atrofia por hipocinesia, uma vez que em um curto período de tempo conseguiram atenuar em 44% a perda de proteínas não-colagenosas do músculo sóleo. Os autores acreditam que contrações excêntricas, durante atividades normais do cotidiano (postura e locomoção), providenciam um potente estímulo para a manutenção do conteúdo protéico muscular. Desta forma, a ausência de contrações excêntricas em condição de hipocinesia pode ser a principal deficiência que inicia a perda de proteínas musculares.
Até mesmo em condição de imobilização por moldes de gesso, a posição em alongamento muscular parece trazer maiores ganhos em termos estruturais em comparação à posição em encurtamento muscular. Quando fibras musculares de animais adultos são imobilizadas em alongamento, ocorre o surgimento de uma maior quantidade de sarcômeros em série e também a redução no comprimento destes como efeito da ação postural. Parece que, como durante o crescimento normal, os sarcômeros séricos são adicionados, principalmente, nas localidades distais do músculo, situação esta explicada se considerarmos o rico suprimento sangüíneo existente nesta região e a necessidade de proteção para o desenvolvimento de uma maior tensão por área de secção transversal. As mudanças em propriedade comprimento-tensão não resultam em alteração da conformação do tecido conjuntivo intramuscular (Williams, Goldspink, 1971; Williams, Goldspink, 1973; Williams, Goldspink, 1978; Alter, 1988; Williams, 1988). Em relação às mudanças ocorrentes na posição imobilizada em encurtamento, nota-se que há uma redução em comprimento das fibras musculares, devido a perdas séricas de sarcômeros, e uma remodelação da coneção dos tecidos conjuntivos intramusculares, propiciando um aumento na rigidez muscular (Williams, 1990). A redução no comprimento das fibras segue-se com diminuição de capacidade do músculo, em decorrência da nova configuração tecidual conjuntiva que favorece um aumento na proporção de colágeno. Esta redução de comprimento não é exclusiva somente quando os músculos são imobilizados numa posição encurtada: um trabalho excessivo em amplitude limitada também poderá levar a uma diminuição do número total de sarcômeros séricos (Williams et al., 1988). A capacidade de produzir tensão máxima sofrerá, desta forma, uma alteração, haja visto que esta é uma adaptação para modificar o comprimento funcional do músculo: numa posição completamente contraída, pouca tensão ativa pode ser desenvolvida (Williams, Goldspink, 1984, Antunes Neto, Vilarta, 1996)
Considerações finais
O presente estudo mostrou as respostas adaptativas positivas advindas do treinamento muscular excêntrico, contrapondo-se aos modelos que apresentam a influência deste tipo de contração no desencadeamento de microlesões ou lesões de maior magnitude, os quais sempre relatam as principais alterações em decorrência de sessões isoladas de ações musculares excêntricas. A sistematização e a constância de execução de exercícios excêntricos, supostamente, providenciam uma condição de estímulo para síntese protéica; a tensão excedente dos elementos elásticos do músculo poderia agir como o estímulo inicial para desencadeamento do evento.
Referências bibliográficas
-
ANTUNES NETO, J. M. F., VILARTA, R. Mecanismos de controle das modificações estruturais do sistema locomotor: construção de modelos explicativos da adaptação postural em situação de imobilização em posições de alongamento e encurtamento musculares. Anais do III Congresso Latino-Americano: Esporte, Educação e Saúde no Movimento Humano, 1996.
-
BENNETT, J. G., STAUBER, W. T. Evaluation and treatment of anterior knee pain using eccentric exercise. Medicine and Science in Sports and Exercise, v. 18, n. 05, p. 526-530, 1986.
-
CARSON, J. A. The regulation of gene expression in hypertrophyng skeletal muscle. Exercise and Sport Sciences Reviews, v. 25, p. 301-320, 1997.
-
CHESLEY, A., MACDOUGALL, J. D., TARNOPOLSKY, M. A. et al. Changes in human muscle protein synthesis after resistance exercise. Journal of Applied Physiology, v. 73, n. 04, p. 1383-1388, 1992.
-
COLLIANDER, E. B., TESCH, P. A. Effects of acute concentric and eccentric exercise on rate of force recovery. Medicine and Science in Sports and Exercise, v. 19, n. 02 (supplement), p. S36, 1987.
-
CROALL, D. E., DEMARTINO, G. N. Calcium-activated neutral protease (calpain) system: structure, function, and regulation. Physiological Reviews, v. 71, n. 03, p. 813-847, 1991.
-
ENOKA, R. M. Neuromechanical basis of kinesiology. Champaign: Human Kinetics, 1994.
-
FRITZ, V. K., STAUBER, W. T. Characterization of muscles injured by forced lengthening. II. Proteoglycans. Medicine and Science in Sports and Exercise, v. 20, n. 04, p. 354-361, 1988.
-
GOLDBERG, A. L., ETLINGER, J. D., GOLDSPINK, D. F. et al. Mechanism of work-induced hypertrofy of skeletal muscle. Medicine and Science in Sport and Exercise, v. 07, n. 04, p. 248-261, 1975.
-
HURME, T., KALIMO, H. Activation of myogenic precursor cells after muscle injury. Medicine and Science in Sports and Exercise, v. 24, n. 02, p. 197-205, 1992.
-
JONES, D. A., RUTHERFORD, O. M. Human muscle strength training: the effects of three different regimes and the nature of the resultant changes. Journal of Physiology, v. 391, p. 1-11, 1987.
-
KIRBY, C. R., RYAN, M. J., BOOTH, F. W. Eccentric exercise training as a countermeasure to non-weight-bearing soleus muscle atrophy. Journal of Applied Physiology, v. 73, n. 05, p. 1894-1899, 1992.
-
KOMI, P. V. Desempenho biomecânico e neuromuscular. Revista Brasileira de Ciências do Esporte, v. 05, n. 02, p. 26-28, 1984.
-
KOMI, P. V. Training of muscle strength and power: interation of neuromotoric, hypertrophic, and mechanical factors. International Journal of Sports Medicine, v. 07, p. 10-15, 1986.
-
MCCOMAS, A. J. Skeletal muscle: form and function. Champaign: Human Kinetics, 1996.
-
MILLWARD, D. J. Protein turnover in skeletal and cardiac muscle during normal growth and hypertrophy. In: WILDENTHAL, K. (org.). Degradative processes in heart abd skeletal muscle: research monographs in cell and tissue physiology. Amsterdam: Elsevier, v. 03, 1980.
-
PATEL, T. J., LIEBER, R. L. Force transmission in skeletal muscle: from actomyosin to external tendons. Exercise and Sport Sciences Reviews, v. 25, p. 321-363, 1997.
-
SALE, D. G. Neural adaptation to resistance training. Medicine and Science in Sports and Exercise, v. 20, n. 05, S135-S145, 1988.
-
SMITH, R. C., RUTHERFORD, O. M. A comparison of eccentric and concentric contractions in strength training in humans. Journal of Physiology, v. 475 P, p. 26 P, 1994.
-
STAUBER, W. T. Eccentric actions of muscles: physiology, injury, and adaptation. Exercise and Sport Science Review, v. 17, p. 157-185, 1989.
-
STAUBER, W. T., FRITZ, V. K., VOGELBACH, D. W. et al. Characterization of muscles injured by forced lengthening. I. Cellular infiltrates. Medicine and Science in Sports and Exercise, v. 20, n. 04, p. 345-353, 1988.
-
TESCH, P. A., DUDLEY, G. A., DUVOISIN, M. R. et al. Force and EMG signal patterns during repeated bouts of concentric or eccentric muscle actions. Acta Physiologica Scandinavica, v. 138, n. 03, p. 263-271, 1990.
-
VIRU, A. Molecular cellular mechanisms of training effects. Journal of Sports Medicine and Physical Fitness, v. 34, n. 04, p. 309-322, 1994.
-
WILLIAMS, P. E. Effect of intermittent stretch in immobilised muscle. Annals of the Rheumatic Diseases, v. 47, p. 1014-1016, 1988.
-
WILLIAMS, P. E. Use of intermittent stretch in the prevention of serial sarcomere loss in immobilised muscle. Annals of the Rheumatic Diseases, v.49, p. 316-317, 1990.
-
WILLIAMS, P. E., GOLDSPINK, G. Longitudinal growth of striated muscle fibres. Journal of Cell Science, v. 09, p. 751-767, 1971.
-
WILLIAMS, P. E., GOLDSPINK, G. The effect of immobilization on the longitudinal growth of strited muscle fibres. Journal of Anatomy, v. 116, n. 01, p. 45-55, 1973.
-
WILLIAMS, P. E., GOLDSPINK, G. Changes in sarcomere length and physiological properties in Immobilised muscle. Journal of Anatomy, v. 127, n. 3, p. 459-468, 1978.
-
WILLIAMS, P. E., GOLDSPINK, G. Connective tissue changes in immobilised muscle. Journal of Anatomy, v. 138, n. 2, p. 343-350, 1984.
-
WILLIAMS, R. S. Frontiers of exercise research: a search for the molecular basis of the exercise training effect in skeletal muscle. In: WILLIAMS, R. S., WALLACE, A. G. (editors). Biological effects of physical activity. Champaign: Human Kinetics Books, 1989.
-
WONG, T. S., BOOTH, F. W. Protein metabolism in rat tibialis anterior muscle after stimulated chronic concentric exercise. Journal of Applied Physiology, v. 69, n. 05, p. 1708-1717, 1990a.
-
WONG, T. S., BOOTH, F. W. Protein metabolism in rat tibialis anterior muscle after stimulated chronic eccentric exercise. Journal of Applied Physiology, v. 69, n. 05, p. 1718-1724, 1990b.
Outros artigos em Portugués
| |
Búsqueda personalizada
|
|---|---|
|
EFDeportes.com, Revista
Digital · Año 16 · N° 164 | Buenos Aires,
Enero de 2012 |
|