
|
|||
|---|---|---|---|
|
|
Cinética de marcadores de estresse oxidativo para avaliação de ‘overreaching’ induzido pelo exercício físico exaustivo La cinética de los marcadores de estrés oxidativo para la evaluación de ‘overreaching’ inducido por el ejercicio física intenso |
|
|
|
*Doutor em Biologia Funcional e Molecular, Instituto de Biologia Departamento de Bioquímica, Universidade Estadual de Campinas, UNICAMP **Bacharel em Esporte, Faculdades Integradas Metropolitanas de Campinas, METROCAMP |
Joaquim Maria Ferreira Antunes Neto* Caio César Donadon** (Brasil) |
|
|
|
Resumo O objetivo deste estudo foi analisar marcadores de estresse em situação de exercício exaustivo de corrida, acompanhando suas cinéticas até 3 horas após o exercício, com o intuito de compreender melhor em qual momento pós exercício havia níveis de estresses oxidativo e fisiológico mais elevados. Participaram do estudo 11 indivíduos do sexo masculino, praticantes de atividades físicas, porém não envolvidos com treinamento sistematizado, com idade entre 20,05±1,97 anos, estatura entre 1,77±0,03 metro e massa corporal entre 77,9±7 quilos. Os exercícios constaram de 2 séries de corridas progressivas na esteira computadorizada (marca IMBRASPORT®), com aumento de velocidade a cada 2 minutos. Os valores de velocidade iniciaram-se em 5 m/s e encerraram-se com o pedido do voluntário (ou se a freqüência cardíaca chegasse numa intensidade próxima a 90% da freqüência cardíaca máxima determinada). Entre uma sessão e outra de exercício exaustivo havia 3 minutos de recuperação. Foram coletados 3 mL de sangue dos voluntários em 5 momentos do experimento: imediatamente antes ao exercício (TA), imediatamente após o exercício (T0h), 1 hora após o exercício (T1h), 2 horas após o exercício (T2h) e 3 horas após o exercício (T3h). Essa condição permitiu gerar uma cinética de estudos dos marcadores, possibilitando compreender em qual momento, após o exercício exaustivo, haveria uma maior condição de estresses oxidativo e fisiológico. Os marcadores estudados foram a enzima antioxidante catalase (CAT) no eritrócito, peroxidação lipídica (método TBARS), creatina quinase (CK- marcador de lesão), ácido úrico (AU) e creatinina (CRE) em amostras de plasma de indivíduos jovens. As amostras foram coletadas antes, imediatamente após, 1 hora, 2 horas e 3 horas após as duas sessões de corrida exaustiva em esteira rolante. Os resultados mostraram que os valores de CAT e TBARS foram significativamente elevados 3 horas após o exercício (p<0.01), com níveis de CK mais significativos 2 horas após o exercício exaustivo (p<0.001). Não houve alterações significativas para os parâmetros fisiológicos estudados (AU e CRE; p>0.05). Concluiu-se que as respostas iniciais de estresse de organismos submetidos a exercícios exaustivos são de origem molecular, sem precipitações de parâmetros fisiológicos em um tempo de 3 horas após a execução do exercício. Portanto, fica evidente que a coleta de sangue para análises clínicas ou de avaliação física não devem ser realizadas imediatamente após o esforço físico, pois se corre o risco de não se observar a maior precipitação de parâmetros de estresse em decorrência da prática exaustiva. Unitermos: Estresse oxidativo. Exercício físico. Overreaching.
|
|||
|
|
EFDeportes.com, Revista Digital. Buenos Aires, Año 16, Nº 162, Noviembre de 2011. http://www.efdeportes.com/ |
|
|
1 / 1
Introdução
A literatura carece de marcadores confiáveis para determinação da fase aguda do overreaching, situação de estresse de característica metabólica, sobretudo no que diz respeito no restabelecimento de substratos energéticos e das enzimas envolvidas nas atividades glicolíticas e neoglicolíticas (Tiidus, 1998). A dificuldade em estabelecer suporte glicogênico para geração pronta de energia nas atividades que envolvem a glicólise anaeróbia ou, bem como, para facilitar recuperação ativa dos atletas durante momentos de intervalo entre um esforço intermitente e outro, é um dos pontos que caracteriza o overreaching (Lehmann et al., 1998).
O objetivo em selecionar os intervalos e cargas de treinamento de modo otimizado encontra-se em assegurar que uma subseqüente sessão de treino coincida com a fase de supercompensação (Bruin et al., 1994), caracterizada como o período de restabelecimento se um determinado substrato metabólico para níveis superiores àqueles encontrados no início da sessão. Desta forma, as adaptações biológicas em longo prazo são caracterizadas pelos efeitos acumulativos das quebras homeostáticas programadas em termos de intensidade, duração, freqüência do próprio estímulo e sua conseqüente resposta adaptativa, o que leva a um estágio superior de performance física (Bompa, 1990). É importante lembrar que a quantidade e a qualidade do estímulo produzam um mesmo aumento adaptação sistêmica do organismo somente quanto este encontra-se preparado para resistir à inibição que pode ser gerada pela intensificação nas cargas de esforço (progressão para o overtraining) (Kuipers, Keizer, 1988).
As nossas análises residem em marcadores bioquímicos e fisiológicos, cujos tempos de alteração/precipitação de parâmetros dependem, sobretudo, no caso das enzimas, de estímulos indutores altamente sensíveis para modificação das propriedades funcionais destas (Antunes Neto et al., 2006). As escolhas por marcadores de estresse oxidativo e de alterações celulares devem-se ao fato de querermos observar como ocorre a modulação da atividade da molécula estudada, o que poderia facilitar o conhecimento das propriedades de regulação alostérica de muitas enzimas e de morfoestruturas submetidas ao estresse oxidativo.
Como um estudo introdutório, buscamos investigar sujeitos saudáveis, não sedentários, porém não praticantes de treinamento sistematizados, com compleição física avaliada como normal. Os marcadores avaliados foram a enzima antioxidante Catalase, no eritrócito (CAT), a enzima Cretina Quinase (CK), as substâncias reativas ao ácido tiobarbitúrico (TBARS) e os níveis plasmáticos de ácido úrico e creatinina, no plasma.
O objetivo deste estudo foi analisar como os marcadores CAT e os níveis de alterações em permeabilidade da membrana celular (CK e TBARS) podem sinalizar o momento de overreaching ou recuperação tecidual, bem como concentrações plasmáticas de ácido úrico e creatinina acompanham a dinâmica de instalação de estresse oxidativo. Uma vez conhecido pouco mais sobre as flutuações destes marcadores, haverá maior facilidade para preparadores físicos e comissões técnico-esportivas, em trabalho conjunto com pesquisadores, em estabelecer fases individualizadas de recuperação e até mesmo de compensação (modulação) da carga de treinamento.
Materiais e métodos
Sujeitos
Participaram do estudo 11 indivíduos do sexo masculino, praticantes de exercícios físicos, porém não envolvidos com treinamento sistematizado, com idade entre 20,05±1,97 anos, estatura entre 1,77±0,03 metro e massa corporal entre 77,9±7 quilos).
Exercício físico
Os exercícios constaram de 2 séries de corridas progressivas na esteira computadorizada (marca IMBRASPORT®), com aumento de velocidade a cada 2 minutos. Os valores de velocidade iniciaram-se em 5 m/s e encerraram-se com o pedido do voluntário (ou se a freqüência cardíaca chegasse numa intensidade próxima a 90% da freqüência cardíaca máxima determinada). Entre uma sessão e outra de exercício exaustivo havia 3 minutos de recuperação. Foram coletados 3 mL de sangue dos voluntários em 5 momentos do experimento: imediatamente antes ao exercício (TA), imediatamente após o exercício (T0h), 1 hora após o exercício (T1h), 2 horas após o exercício (T2h) e 3 horas após o exercício (T3h). Essa condição permitiu gerar uma cinética de estudos dos marcadores, possibilitando compreender em qual momento, após o exercício exaustivo, haveria uma maior condição de estresses oxidativo e fisiológico.
Amostras de plasma e preparação de hemolisados
Amostras de sangue foram coletadas da veia hepática, utilizando seringas heparinizadas. O plasma foi separado das hemácias por centrifugação a 1.000g por 10 minutos e então transferidas para um tubo novo e armazenadas a –80ºC até a análise. Os eritrócitos foram lisados com o dobro de água destilada (1:1, v/v) e armazenados a –80ºC, até a determinação da atividade de catalase (CAT) e glutationa redutase (GR). A concentração de Hb foi determinada pelo método Drabkin.
Dosagens enzimáticas
-
Creatina Quinase:
Catalase:
Dosagem de Hemoglobina no Hemolisado.
Dosagens de Parâmetros Metabólicos.
Peroxidação Lipídica (TBARS).
Análises estatísticas
Utilizamos o software GraphPad Instat® (San Diego-CA) para conduzir as análises estatísticas. O teste apropriado foi “one way” ANOVA para amostras pareadas e o teste Tukey foi adotado como pós teste.
Resultados
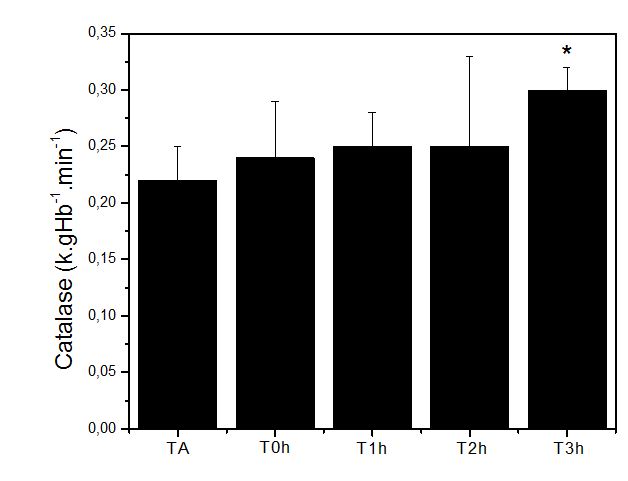
Figura 1. Níveis no hemolisado de Catalase (k.gHb-1.min-1) obtidos antes (TA), imediatamente após (T0h), uma hora após (T1h), duas horas após
(T2h) e 3 horas após (T3h) a execução do protocolo de exercício exaustivo e não acostumado em esteira rolante. Onde: * = p<0.05 em relação à TA
Os resultados mostram que houve maior atividade significativa (p<0.05) da enzima CAT após 3 horas de execução do exercício exaustivo, em relação ao momento T0. Essa evidência demonstra grande capacidade de regulação desta enzima antioxidante em condição de requerimento exaustivo das exigências metabólicas.
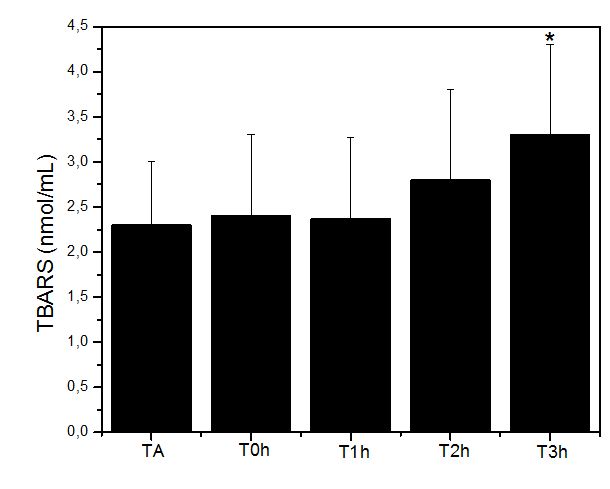
Figura 2. Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS; nmo/mL) obtidos antes (TA), imediatamente após (T0h), uma hora após (T1h), duas
horas após (T2h) e 3 horas após (T3h) a execução do protocolo de exercício exaustivo e não acostumado em esteira rolante. Onde: * = p<0.05 em relação à TA
Da mesma forma observada para a atividade antioxidante (CAT), o parâmetro de alteração de permeabilidade lipídica de membrana (TBARS) foi afetado mais significativamente 3 horas após o exercício exaustivo, o que evidencia a instalação de um quadro de estresse oxidativo.
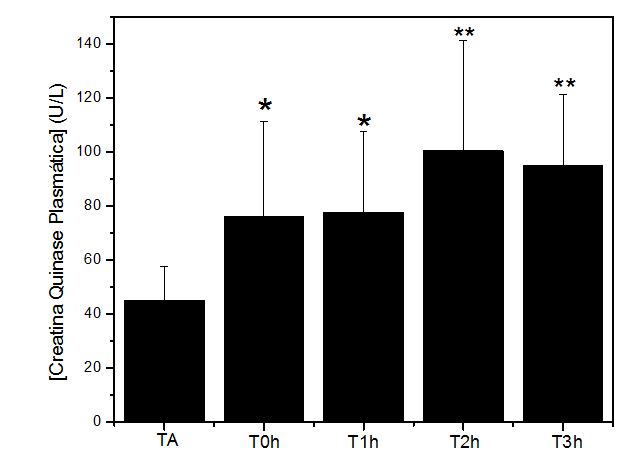
Figura 3. Níveis plasmáticos da enzima Creatina Quinase (U/L) obtidos antes (TA), imediatamente após (T0h), uma hora após (T1h), duas horas após (T2h) e 3 horas após (T3h) a
execução do protocolo de exercício exaustivo e não acostumado em esteira rolante. Onde: * = p<0.05 em relação a TA; ** = p<0.01 em relação a TA e p<0.05 em relação a T0h e T1h
A precipitação da concentração plasmática de CK (U/L) inicia-se imediatamente após o exercício exaustivo, possivelmente pelo aumento das microlesões induzidas pelos estresses mecânico e metabólico. A maior concentração média de CK no plasma ocorreu 2 horas após o exercício, 1 hora antes do observado para CAT e TBARS. Tal condição sugere uma estratégia de regulação para atenuação do estresse oxidativo aparentemente instalado.
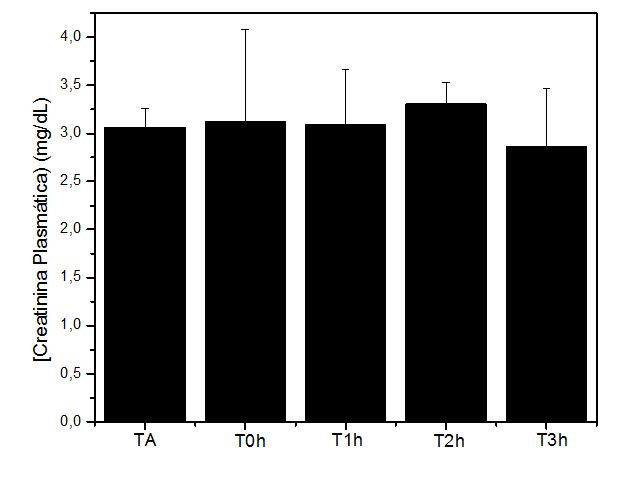
Figura 4. Níveis plasmáticos de Creatinina (mg/dL) obtidos antes (TA), imediatamente após (T0h), uma hora após (T1h), duas horas após (T2h) e 3 horas
após (T3h) a execução do protocolo de exercício exaustivo e não acostumado em esteira rolante. Não houve diferença estatística entre as análises
Os valores para parâmetros metabólicos, tal como creatinina (mg/dL), não demonstraram alterações significativas, sugerindo que as estratégias moleculares (CAT, CK e TBARS) antecipam os eventos de mudanças na célula.
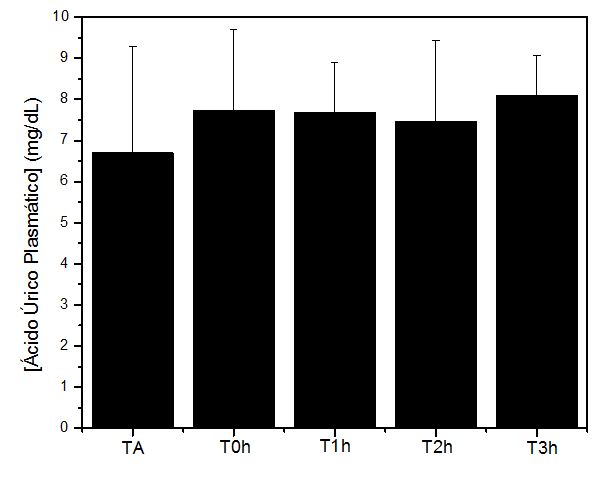
Figura 5. Níveis no plasmáticos de ácido úrico (mg/dL) obtidos antes (TA), imediatamente após (T0h), uma hora após (T1h), duas horas após (T2h)
e 3 horas após (T3h) a execução do protocolo de exercício exaustivo e não acostumado em esteira rolante. Onde: * = p<0.05 em relação à TA
Os valores para parâmetros metabólicos, tal como ácido úrico (mg/dL), não demonstraram alterações significativas, sugerindo que as estratégias moleculares (CAT, CK e TBARS) antecipam os eventos de mudanças na célula
Discussão
O início do overtraining é conhecido na literatura como overreaching e se instala quando o repouso entre o próximo treinamento ou competição é insuficiente (Bruin et al., 1994, Tiidus, 1998; Lehmann et al., 1998). Embora esta condição também induza fadiga prematura, pois a recuperação é incompleta, pode ser facilmente revertida com um ou dois dias de pouco ou nenhum treino. Entretanto, como pouco é conhecido em relação à quantidade ótima de treinamento e os fatores que influenciam a recuperação e supercompensação, ainda hoje existe um componente muito grande de empirismo na aplicação dos métodos de treinamento, que são na sua grande maioria empregados sem muita base científica (Lazarim et al., 2009). Ou seja, os atletas normalmente são submetidos a cargas de esforço muito altas, com pouco tempo de recuperação, para não se pecar pela falta de estímulo, sendo impossível prever se a carga de esforço exigida da equipe está excedendo o limite individual de estresse que o organismo pode suportar. Tudo isso torna o limiar entre um treinamento ideal e o overtraining muito tênue. O maior problema é que ainda não existem descritos na literatura marcadores completamente confiáveis, nem de adaptação, nem de overreaching, que ajudem a diagnosticar esse limite individual de estresse que impeça a instalação do overtraining. Além disso, a literatura é carente de protocolos de exercícios que se preocupem em estabelecer esse quadro de desbalanço entre exercício/repouso em animais de laboratório, o que também dificulta o estudo da sensibilidade de biomarcadores nessa situação (Antunes Neto et al., 2008).
Uma via de sinalização aparentemente envolvida com esses dois processos antagônicos, adaptação e overtraining, muito discutida na literatura, envolve a alteração no estado redox intracelular, induzida pela elevação na produção de espécies reativas de oxigênio (EROs) que acompanha o aumento da atividade contrátil muscular (Smolka et al., 2000; Reid, 2001).
As EROs são espécies químicas independentes, que possuem um ou mais elétrons desemparelhados. Isto implica em uma grande instabilidade e, freqüentemente, elevada reatividade. Como necessitam completar seus pares de elétrons para se estabilizarem, comportam-se ou como receptores (oxidantes) ou como doadores (redutores) de elétrons. Nesta definição incluem-se os átomos de hidrogênio, vários íons metálicos de transição (ferro e cobre) e o oxigênio molecular (Halliwell, Gutteridge, 1989). Segundo Schneider e Oliveira (2004) as EROs são produzidas naturalmente em nosso organismo através de processos metabólicos oxidativos, sendo importantes em situações de necessidade de ativação do sistema imunológico. Por outro lado, a produção excessiva de EROs pode levar a uma situação de estresse oxidativo com subseqüentes efeitos prejudiciais ao organismo, tais como peroxidação lipídica, oxidação de proteínas, agressão a carboidratos e ao ácido desoxirribonucléico (DNA).
Nosso estudo mostrou que uma única sessão de exercício exaustivo de corrida, realizada por sujeitos saudáveis ativos, pode modificar a homeostasia celular. O evento inicial para a instalação de um quadro de estresse oxidativo foi o aumento da concentração plasmática da enzima CK (Figura 3). Estudos prévios mostraram que a liberação de CK no plasma tem relação direta com os mecanismos de microlesões celulares, decorrentes de distúrbios de ordem mecânica e/ou metabólica. A causa metabólica de microlesão na célula muscular parece estar envolvida com uma produção inadequada de ATP em relação à sua demanda, podendo resultar num processo isquêmico que favorece a degradação de estruturas protéicas, ocasionando um quadro de prejuízo citoesquelético (Rubin et al., 1996). Além do mais, o exercício físico pode ser uma fonte indutora para produção de EROs, conduzindo a um aumento de distúrbios celulares, principalmente de proteínas e lipídios de membranas (Antunes Neto et al., 2005). Considerando alterações de ordem mecânica, inúmeras evidências apontam que exercícios que se utilizam basicamente de contrações musculares excêntricas tendem a desencadear um maior número de respostas lesivas no meio celular, tal como distúrbios nas linhas Z e bandas A dos sarcômeros e ruptura do retículo sarcoplasmático (Lieber et al., 1996; Teague, Schwane, 1995; Nosaka, Clarkson, 1994).
No modelo de estudo utilizado por nós – corrida em esteira rolante – a transição da fase excêntrica para a fase concêntrica do movimento ocorre de forma vigorosa, com alto requerimento metabólico. Uma vez tal quadro instalado, há uma concomitante produção de EROs e um desencadeamento de microlesões celulares, possibilitando que haja distúrbios severos na membrana celular e, desta forma, aumento de oxidação dos lipídios (Figura 2). A estratégia de regulação celular defronte tal situação, para evitar que haja a prevalência do ataque oxidativo, é o aumento da atividade do sistema antioxidante. Nossos dados mostram que a CAT (Figura 1) eleva sua capacidade funcional simultaneamente à precipitação de TBARS (Figura 2) e posteriormente ao início do aumento de permeabilidade da membrana celular, condição ilustrada pelos valores de CK (Figura 3). A função da CAT é detoxificar o peróxido de hidrogênio (H2O2) gerado ao final da cadeia transportadora de elétrons, a qual tem como objetivo gerar ATPs. Torna-se importante evitar o aumento de concentração de H2O2 celular, pois, apesar deste composto ser considerado uma espécie radicalar fraca, ele possui a propriedade de atravessar facilmente membranas celulares e a união com um elétron proveniente de metais de transição, como Fe2+ ou Cu+, poderia dar origem ao radical hidroxila (˙OH), uma das espécies radicalares existentes mais reativas. Esta condição permitiria o aumento dos níveis de TBARS (Figura 2), refletindo na desintegração lipídica celular e no aumento do estresse oxidativo.
Concluindo, o aumento significativo da atividade de CAT, mesmo com elevação simultânea dos níveis de CK e TBARS, permite considerar que o organismo de sujeitos jovens e saudáveis, mesmo não acostumados com a execução de exercícios exaustivos nas suas rotinas de atividades físicas, consegue estabelecer uma estratégia de atenuação de estresse oxidativo. Os valores não significativos obtidos para creatinina (Figura 4) e ácido úrico (Figura 5), utilizados como indicadores de catabolismo muscular, sugerem que a condição de distúrbio celular obtida pode gerar uma condição adaptativa positiva, caso os intervalos de recuperação – mínimo de 3 horas – sejam respeitados entre um ciclo de exercícios exaustivos e outro. Evidencia-se que uma estratégia de supercompensação seria satisfatória quando um estímulo estressor fosse aplicado no momento de maior atividade antioxidante celular, evitando, portanto, que danos secundários lesivos à célula sejam elevados.
Referências bibliográficas
-
AEBI, H. Catalase. In: Methods in enzymology. Packer, L (editor). Florida: Academic Press, 105, p. 121-126, 1984.
-
ANTUNES NETO J. M. F., PEREIRA-DA-SILVA, L., MACEDO, D. V. Biomarcadores de estresse oxidativo: novas possibilidades de monitoramento em treinamento físico. Revista Brasileira de Ciência e Movimento, v. 13, n. 02, p. 73-79, 2005.
-
ANTUNES NETO, J. M. F., PEREIRA-DA-SILVA, L., MACEDO, D. V. Heat shock proteins as oxidative stress markers in rats submitted to exhaustive intermittent running training. Brazilian Journal of Biomotricity, v.02, n. 03, p. 160-175, 2008.
-
ANTUNES NETO J. M. F., TOYAMA M. H., CARNEIRO E. M., BOSCHERO A. C., PEREIRA-DA-SILVA L., MACEDO D. V. Circulating leukocyte heat shock protein 70(hsp70) and oxidative stress markers in rats after a bout of exhaustive exercise. International Journal on the Biology of Stress, v. 09. n. 02. p. 107–115, 2006.
-
BEUTLER, E. Red cell metabolism. In: A manual of biochemical methods. London: Grune & Stratton Publishers, 1975.
-
BOMPA, T. O. Theory and methodology of training: to key to athletic performance. Dubuque: Kendall/Hunt, 1990.
-
BRUIN, G. KUIPERS, H., KEIZER, H. A., VANDERVUSSE, G. J. Adaptation and Overtraining in horses subjected to increasing training loads. Journal of Applied Physiology, v. 76, n. 05, p. 1908-1913, 1994.
-
HALLIWELL, B.; GUTTERIDGE, J. M. Free radicals in biology and medicine. Oxford: Clarendon Press, 1989.
-
KUIPERS, H., KEIZER, H. A. Overtraining in elite athletes: review and directions for the future. Sports Medicine, v. 06, p. 79-92, 1988.
-
LAZARIM, F. L., ANTUNES NETO, J. M. F., SILVA, F. O. C., NUNES, L. A. S., BASSANI-CAMERON, A., CAMERON, L. C., ALVES, A. A., BRENZIKOFER, R., MACEDO, D. V. The upper values of plasma creatine kinase of professional soccer players during the Brazilian National Championship. Journal of Science and Medicine in Sport, v. 12, p. 85-90, 2009.
-
LEHMANN, M., FOSTER, C., DICKHUTH, H. H., GASTMANN, U. Autonomic imbalance hypothesis and overtraining syndrome. Medicine and Science in Sports and Exercise, v. 30, n. 07, p. 1140-1148, 1998.
-
LIEBER, R. L.; THORNELL, L. E.; FRIDÉN, J. Muscle cytoskeletal disruption occurs within the first 15 min of cyclic eccentric contraction. Journal of Applied Physiology, v. 80, n. 1, p. 278-284, 1996.
-
NOSAKA, K.; CLARKSON, P. M. Effect of eccentric exercise on plasma enzyme activities previously elevated by eccentric exercise. European Journal of Applied Physiology and Occupational Physiology, v. 69, n. 6, p. 492-497, 1994.
-
REID, M. B. Redox Modulation of skeletal muscle contraction: what we know and what we don’t. Journal of Applied Physiology, v. 90, p. 724-731, 2001.
-
RUBIN, B. B., ROMASCHIN, A., WALKER, P. M. Mechanisms of postischemic injury in skeletal muscle: intervention strategies. Journal of Applied Physiology, v. 80, n. 2, p. 369-387, 1996.
-
SCHNEIDER C. D., OLIVEIRA A. R. Radicais livres de oxigênio e exercício: mecanismos de formação e adaptação ao treinamento físico. Revista Brasileira de Medicina do Esporte, v. 10, n.04, p. 01-10, 2004.
-
SMOLKA, M. B., ZOPPI, C. C., ALVES, A. A., SILVEIRA, L. R., MARANGONI, S., PEREIRA-DA-SILVA, L., NOVELLO, J. C., MACEDO, D. V. HSP72 as a complementary protection against oxidative stress induced by exercise in the soleus muscle of rats. American Journal of Physiology, v. 279, p. R1539-R1545, 2000.
-
TEAGUE, B. N., SCHWANE, J. A. Effect of intermittent eccentric contractions on symptoms of muscle microinjury. Medicine and Science in Sports and Exercise, v. 27, n. 10, p. 1378-1384, 1995.
-
TIIDUS, P. M. Radical species in inflammation and overtraining. Canadian Journal of Physiology and Pharmacology, v. 76, p. 533-538, 1998.
-
YAGI, K. A. A simple fluorimetric assay for lipoperoxide in blood plasma. Biochemistry and Medicine, v. 15, p. 212-216, 1976.
Outros artigos em Portugués
| |

Búsqueda personalizada
|
|---|---|
|
EFDeportes.com, Revista
Digital · Año 16 · N° 162 | Buenos Aires,
Noviembre de 2011 |
|