1.
Introducción
Aunque la mayoría de los seres humanos viven a nivel del mar o a poca altura,
40 millones de personas viven a más de 3.000 metros de altura y unos 140
millones viven a más de 2.500 metros. Sin embargo, estos no son los datos que
nos llaman más la atención sino el que en el pueblo minero de La Rinconada
(Perú) viven 7.000 personas a 5.100 metros de altitud (Calbet,
2006).
El estudio de las respuestas agudas y crónicas a la hipoxia ha llamado la
atención de numerosos fisiólogos desde finales del siglo XIX. Pero es en 1968
(nominación de la ciudad de México sede de los juegos olímpicos) cuando se
produce el punto de inflexión en cuanto a estudios científicos que se han
mantenido hasta nuestros días sobre la tolerancia de los seres humanos a la
falta de oxígeno y a los efectos perjudiciales que puede traer consigo.
2.
Diferencias entre
el ambiente en altura y a nivel del mar.
La hipoxia es el principal factor responsable de la mayoría de los efectos
nocivos de la altura sobre la salud (Bert, 1898, citado por Calbet, 2006). No
obstante, no debemos dejar de lado a otros factores como el frío, la
deshidratación, la irradiación solar y las radiaciones ionizantes porque
también son responsables del deterioro de la salud del ser humano en altura.
Exponencialmente, la presión atmosférica desciende en la medida que va
aumentando la altura, pero tenemos que tener en cuenta dos factores (a una misma
altura): el primero es la temperatura, ya que va a descender menos la presión
cuanto mayor sea la temperatura. El segundo factor es la latitud, ya que cuanto
más nos alejemos del ecuador la presión va a ser menor, es decir, a mayor
latitud vamos a tener menos presión.
Con la altura, la humedad ambiental va a descender porque, debido al
descenso de la temperatura, va a haber menos moléculas de agua en el aire
(humedad relativa). Ese hecho produce una pérdida importante de agua corporal
porque, al inspirar el aire frío, este aire se calienta dentro de nuestros
pulmones y, por lo tanto, admite más vapor de agua procedente de nuestro cuerpo
que al exhalarse se pierde a temperatura corporal.
La radiación solar y sus efectos dañinos van a verse aumentados con la
altura por tres aspectos principales: El primero es que la capa atmosférica de
aire que nos protege de esta radiación es más delgada por lo que vamos a tener
menos protección. El segundo es la menor cantidad de moléculas de vapor de
agua existentes, las cuales absorben la radiación solar. Y, por último, la
existencia de nieve aumenta la radiación, ya que la refleja hacia nuestro
cuerpo.
3.
Respuestas agudas a la hipoxia
3.1.
Respuesta ventilatoria a la hipoxia aguda
La presión parcial arterial de oxígeno en sangre (PaO2)
es sólo ligeramente inferior a la presión parcial alveolar de oxígeno (PAO2).
Pero la PAO2
(incluso durante la hiperventilación máxima), es siempre menor que la presión
inspiratoria lo que produce una situación denominada hipoxemia (la PaO2
es inferior en altura). La hipoxemia, por lo tanto, compromete la distribución
de oxígeno a los tejidos (Calbet, 2006).
En el momento que el organismo se somete a la hipoxia, se ponen en marcha una
serie de cambios fisiológicos que intentan devolver el equilibrio, es decir,
restablecer los niveles de oxígeno de la sangre arterial. Pues bien,
automáticamente tendrá lugar un aumento de la ventilación (hiperventilación)
que se debe a la estimulación que la hipoxia produce en los quimiorreceptores
periféricos (los carotídeos, principalmente).
Durante el ejercicio en hipoxia esta respuesta ventilatoria al esfuerzo se
exagera, por lo que, a la misma intensidad absoluta de esfuerzo, el cociente
equivalente del oxígeno (VE/VO2)
es mucho mayor en altura que a nivel del mar. Aquí tenemos una de las causas
por las que en altura, para realizar un esfuerzo a la misma intensidad, hay que
gastar más energía que a nivel del mar (por un mayor trabajo respiratorio).
La hiperventilación provoca también un descenso de la presión parcial
arterial de CO2
(PaCO2)
y, como consecuencia, se produce un ligero aumento de pH sanguíneo (alcalosis
respiratoria) (Calbet, 2006). A los pocos días, el pH sanguíneo vuelve a
aumentar pero siempre dejando cierto grado de alcalosis respiratoria debido a la
secreción de bicarbonato por parte de los riñones. Este grado de alcalosis
permite la fijación de oxígeno a la hemoglobina en el alveolo lo que hace que
se desplace hacia la izquierda la curva de disociación de la hemoglobina. Esto
lo demuestran Calbet y cols. (2003) en su estudio donde descubren que, con este
desplazamiento hacia la izquierda, la hemoglobina es capaz de captar más
oxígeno para llevar a los tejidos, compensando así, el descenso de la presión
arterial de oxígeno y, por tanto, el descenso del VO2
MAX
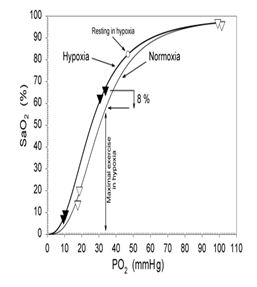
Figura
1. Efectos de la alcalosis respiratoria sobre la curva de disociación de la
hemoglobina. (Calbet JA, Boushel R, Radegran G, Sondergaard H,
Wagner
PD, Saltin B. Determinants of maximal oxygen uptake in severe acute hypoxia. Am
J Physiol Regul Integr Comp Physiol. 2003a; 284: R 291-303)
Por lo tanto, durante un esfuerzo máximo en hipoxia, se produce un descenso de
la concentración arterial de oxígeno que se debe a dos factores principales
que son la disminución de la PiO2
y el empeoramiento del intercambio de gases pulmonar.
El grado de desaturación de la hemoglobina es proporcional al VO2max,
demostrando así que los deportistas de élite presentan un mayor grado de
desaturación que las personas no entrenadas (Lawler y co1s. 1988).
3.2.
Respuesta cardiovascular a la hipoxia aguda
Para evitar el desequilibrio que produce el descenso de la concentración
arterial de oxígeno y hacer que llegue el oxígeno que necesitan los tejidos,
se produce un aumento del flujo de sangre hacia los mismos. Esto se realiza,
como hemos dicho antes, mediante la activación al detectar hipoxia de los
quimiorreceptores periféricos de los cuerpos carotídeos que provocan una serie
de mecanismos fisiológicos que dan como resultado un aumento de la actividad
del sistema nervioso simpático que se traduce en un aumento de la frecuencia
cardíaca, tanto en reposo como a una misma intensidad absoluta de esfuerzo.
Otro efecto de la actividad simpática es evitar que la hipoxia produzca una
caída de la presión arterial con su efecto vasodilatador.
Si nos referimos a la frecuencia cardíaca máxima y al gasto cardíaco máximo,
ambo disminuyen respectivamente en hipoxia aguda. La frecuencia cardíaca
máxima disminuye un latido por cada 130 metros de altitud partir de los 3.000
metros de altura (Lundby y cols. 2001). Como consecuencia del descenso de la
frecuencia cardíaca máxima, es inevitable que descienda el gasto cardíaco
máximo ante un mismo volumen sistólico. No se conoce con exactitud la razón
por la que se producen estas disminuciones pero parece ser que se le puede
atribuir al efecto de la hipoxia sobre el sistema nervioso central (Calbet y
cols. 2003).
En hipoxia aguda aumenta la presión arterial en la arteria pulmonar por lo que
se produce hipertensión cuya magnitud depende del grado de hipoxia existente.
Esta hipertensión que se da en hipoxia se ha relacionado con el empeoramiento
del intercambio de gases alveolar. Se ha demostrado que el sidenafil (la viagra)
reduce ligeramente la hipertensión en la arteria pulmonar en hipoxia en reposo
y durante el ejercicio llegando a la conclusión de que esta sustancia aumenta
el rendimiento en condiciones de hipoxia (Ghofrani y cols. 2004) y Richalet y
cols. 2004)
Par sintetizarlo todo, Calbet y cols. (2003) resumieron los tres mecanismos
responsables de la disminución del consumo máximo de oxígeno en hipoxia aguda
severa y son la reducción de la PiO2,
la disminución de la difusión de gases en el pulmón y la reducción de la
frecuencia cardíaca máxima.
3.3.
Respuestas hematológicas a la hipoxia aguda
El aparato yuxtaglomerular de los riñones detecta el descenso de la
concentración de oxígeno en la sangre y hace que se estimule la segregación
de eritropoyetina (EPO). La EPO estimulará la producción de glóbulos rojos en
la médula ósea de los huesos largos, lo que aumentará la capacidad de
transporte de oxígeno. El factor inducible por hipoxia (HIF) es el encargado de
activar la expresión del gen que codifica la EPO (ARNm EPO).
Cuando nos sometemos a una hipoxia aguda, la liberación de EPO depende del
tiempo que estamos sometidos y del grado de la misma. Eckardt y cols. (1989),
exponiendo a seis sujetos a diferentes alturas, demostraron que son necesarios
de 90 a 120 minutos de exposición para que aumente la concentración de EPO en
sangre y que, una vez terminada esta exposición a hipoxia, la EPO va siendo
eliminada poco a poco de la circulación (la vida media de la hormona es de 5,2
horas). Por debajo de los 2.000 metros aumentan muy poco los niveles de EPO,
mientras que por encima de 2.500 metros este aumento es considerablemente mayor
(Ge y cols. 2002). Al cabo de una exposición a hipoxia de 48 horas, la
concentración de EPO en sangre baja progresivamente hasta alcanzar los niveles
normales en días o 2-3 semanas, dependiendo de la altitud y del individuo
(Milledge y Cotes, 1985).
4.
Aclimatación a la altura
4.1.
Volumen plasmático y agua corporal
En las primeras 24-48 horas en hipoxia, disminuye el volumen plasmático debido
a la diuresis, a la pérdida de agua por sudoración y por ventilación y al
desplazamiento de agua desde el espacio vascular a los espacios extravascular e
intracelular. Éste movimiento de agua (y de proteínas) se debe a la acción
del péptido atrial natriurético que actúa como vasodilatador y aumenta la
permeabilidad capilar (Calbet, 2006).
Esta disminución del volumen plasmático es proporcional al grado de hipoxia y
hace que disminuya el volumen sanguíneo total que se traduce en un 50% de la
pérdida de volumen plasmático total (Sánchez y cols 1970). Las personas que
viven en altura permanentemente tienen un volumen plasmático menor en un 27% a
los residentes a nivel del mar.
La disminución del volumen plasmático tiene la ventaja de permitir aumentar el
hematocrito (sin contar con la EPO). Mientras que en altura pasa esto, a la
vuelta a nivel del mar pasa lo contrario, es decir, se produce una expansión
del volumen plasmático, por lo que el hematocrito vuelve a sus volúmenes
anteriores rápidamente (24-48 horas), aunque haya habido un aumento de
eritrocitos.
4.2.
Ventilación pulmonar e intercambio de gases pulmonar y equilibrio ácido-base
La aclimatación a la hipoxia consiste en aumentar la ventilación en reposo y,
en seres humanos, se lleva a cabo en 10 días aproximadamente (Dempsey y Foster,
1982). Con la exposición muy prolongada a la hipoxia podemos observar un
descenso de la sensibilidad a la misma y de la ventilación en reposo (aunque
siempre sigue siendo superior a la del nivel del mar).
Con la exposición crónica a la hipoxia, los gases sanguíneos sufren cambios
que van a depender del grado de hipoxia (altitud). La PaO2
va a disminuir aunque menos que la PiO2
por el efecto compensatorio del aumento de la ventilación. Mientras, la PaCO2
también va a disminuir de forma importante debido al aumento de la ventilación
dando lugar a una alcalosis respiratoria compensada o no compensada (más de
5.000 metros usualmente).
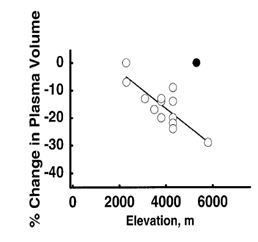
Figura
2. Relación entre la altitud y la pérdida de volumen plasmático. (Swaka
MN, Convertino VA, Eichner ER, Schnieder SM, Young AJ. Blood volumen: importance
and adaptations
to
exercice training, enviromental stresses, and adaptations to exercice training,
environmental stresses, and trauma/sickness. Med Sci Sports Exerc, 2000;
32:332-348).
El aumento de la concentración de hemoglobina producido por la
hemoconcentración que puede deberse a la pérdida de plasma (al principio) o
por la estimulación de la eritropoyesis (a largo plazo) permiten aumentar el
contenido de oxígeno de la sangre arterial en reposo.
Con la aclimatación a alturas moderadas, la respuesta ventilatoria al esfuerzo
máximo se vuelve a situar cerca de los niveles a nivel del mar. Pero en hipoxia
severa, aumenta la respuesta ventilatoria a un esfuerzo máximo. Con la
aclimatación, la PAO2
es superior al mismo nivel de ventilación (mejora la eficiencia ventilatoria).
Esto lo demostraron Lundby y cols. (2004) cuando estudiaron a seis sujetos a
nivel del mar, en hipoxia aguda y después de 2 y 8 semanas de aclimatación a
4.100 metros de altura. La mejora de la oxigenación arterial (PaO2)
durante el esfuerzo máximo es el resultado de una mejora en la ventilación
alveolar y en la PAO2.
Pero, a alturas más elevadas, la mejora de la PaO2
es debida a dos mecanismos principales que son el aumento de la PAO2
y de una mejora en el intercambio de gases como reflejan (Calbet y cols. 2003).
La saturación de la hemoglobina con oxígeno (SaO2)
depende principalmente de la PaO2
y de la afinidad de la hemoglobina por el oxígeno. El aumento de esta afinidad
es debido principalmente a que con el aumento de la ventilación se produce
alcalosis la cual estimula la producción de 2,3-difosfoglicerato (2-3 DPG) en
los eritrocitos. Este producto disminuye la afinidad de la hemoglobina por el
oxígeno, es decir, desplaza la curva de disociación de la hemoglobina hacia la
derecha. Por lo tanto se producen dos movimientos antagónicos: la alcalosis
respiratoria que desplaza la curva de disociación de la hemoglobina hacia la
izquierda y el aumento de 2-3 DPG que la desplaza hacia la derecha. El resultado
de todo esto es que en reposo, la afinidad de la hemoglobina por el oxígeno en
el tejido en altura es similar a la que se da a nivel del mar y que, sin
embargo, a grandes alturas y en reposo, la curva se desplaza ligeramente hacia
la izquierda para facilitar la captación de oxígeno en el alveolo.
4.3.
Cambios en el aparato circulatorio
Los principales cambios que tienen lugar en el aparato circulatorio como
consecuencia de la aclimatación a la altura son el aumento de la actividad
simpática y parasimpática, aumento de la presión arterial sistémica y
pulmonar en reposo, aumento de la frecuencia cardíaca en reposo, disminución
del gasto cardíaco máximo, disminución de la frecuencia cardíaca máxima y
aumento de la de la densidad capilar en la musculatura esquelética. El aumento
de la actividad simpática provocaría un aumento de la frecuencia cardíaca en
reposo hasta 140-150 latidos/minuto si no existiera el aumento de la actividad
parasimpática que la lleva a latidos/minuto muy cercanos a los del nivel del
mar. Sin este aumento de la actividad parasimpática, en pocos días se
produciría una insuficiencia cardiaca provocada al efecto negativo de la
taquicardia sobre el miocardio (Calbet, 2006).
A 4.000 metros de altura, el descenso del gasto cardiaco máximo se asocia a la
aclimatación. Esto no se debe a una disminución de la frecuencia cardíaca
como se creía anteriormente sino que, como demostraron Boushel y cols. (2001)
en un estudio que realizaron con siete sujetos sanos durante nueve semanas, que,
aunque esta frecuencia cardíaca se restaure, el gasto cardíaco máximo sigue
disminuido. El gasto cardíaco no disminuye debido exclusivamente al descenso
del volumen plasmático, al aumento del hematocrito ni a la hipertensión
pulmonar (Calbet y cols. 2003 y 2004 y Richalet y cols. 2004).
4.4.
Aclimatación a la altura: cambios hematológicos
Tras pasar de uno a tres meses en alturas superiores a los 3.000 metros se puede
observar como incrementa progresivamente la masa eritrocitaria. Reynafarje y
cols. (1959) estudiaron a diez militares destinados a 4.509 metros en Perú
durante un año y, al cabo de un mes, detectaron un ligero aumento de la masa
eritrocitaria que siguió aumentando hasta los ocho meses (a los ocho meses la
masa eritrocitaria había aumentado un 50%). Sin embargo, después de dos meses
de permanencia a 4.300 metros, Hannon y cols. (1969) no observaron aumento en la
masa eritrocitaria (incluso suplementando a los sujetos con hierro). En general,
la masa eritrocitaria no empieza a aumentar hasta que la PaO2
cae por debajo de 70 mmHg (es equivalente a alturas de 2.500-3.000 metros) (Weil
y cols. 1968).
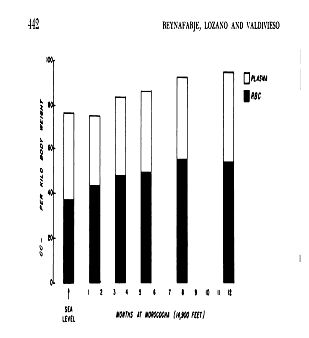
Figura
3. Cambios en el volumen plasmático (blanco) y en la masa eritrocitaria
(negro) durante la permanencia prolongada a unos 4.500 m de altura.
(Reynafarje
C, Lozano R, Valdivieso J. The polycythemia of high altitude: iron metabolism
and related aspects. Blood. 1959; 14: 433-55).
4.5.
Aclimatación a la altura: adaptaciones neuroendocrinas
Especialmente a altitudes superiores a 5.000 metros se produce un aumento de las
catecolaminas circulantes. Durante las primeras semanas, aumentan la ACTH, el
cortisol y las hormonas tiroideas (vuelven a los niveles propios del nivel del
mar hacia la tercera-cuarta semana de estancia en altura). La mayor parte de la
respuesta hormonal que tiene lugar en los primeros días es para fomentar la
diuresis y la natriuresis (eliminación de sodio a través de la orina). En la
primera semana de exposición a la hipoxia, se produce una disminución de la
actividad del sistema renina-angiotensina-aldosterona que al cabo de unas
semanas vuelve a la actividad normal (Zaccaria y cols, 1998).
5.
Capacidad de rendimiento físico en altura
Dependiendo de las características del entrenamiento y de la altura en la que
se realice, el rendimiento en altura puede verse aumentado, disminuido o sin
variaciones. Por ejemplo, se obtienen mejoras en carreras de atletismo de menos
de dos minutos mientras que en las superiores a dos minutos se empeora el
rendimiento (Fulco y cols. 1998). La razón por la que no empeoran las carreras
de menos de dos minutos es porque en este tipo de carreras se utilizan
mayoritariamente mecanismos anaeróbicos que son menos sensibles a la hipoxia
que los aeróbicos. Por lo tanto, la hipoxia no afecta a los mecanismo
anaeróbicos pero si a los aeróbicos. Esta mejora en las carreras de corta
duración también se debe, en segundo lugar, a que en altura la resistencia que
produce el aire es menor porque con la altitud disminuye su densidad. Este hecho
produce mejoras del rendimiento en pruebas explosivas y de sprint como son las
carreras de 100 o 400 metros, saltos, lanzamientos, etc.
Podemos sintetizar todo lo anterior diciendo que solo obtendremos mejoras en
altura en aquellas disciplinas deportivas en las que el rozamiento del aire sea
determinante. Sin embargo, en deportes como el ciclismo se puede mejorar en
altura si se habla del ahorro energético al ofrecer menos resistencia el aire y
este ahorro energético es mayor cuanto mayor sea la velocidad incluso pudiendo
llegar a contrarrestar los deterioros del rendimiento en altura a grandes
velocidades (Calbet, 2009).
Con la aclimatación mejora notablemente el rendimiento en pruebas de
resistencia efectuadas en altura. Siguiendo a Maher y cols. (1974), se puede
mejorar en un 45%.
6.
Efectos de la altitud sobre el VO2MAX
En hipoxia se produce una disminución de la saturación máxima que puede
alcanzar la hemoglobina durante el ejercicio por lo que se compromete el
suministro de oxígeno. Como consecuencia de esto, disminuye el rendimiento en
pruebas en las que se utilice fundamentalmente el metabolismo aeróbico porque,
en altura, se pude mantener una misma intensidad relativa de esfuerzo durante
menos tiempo y la cantidad absoluta máxima del VO2
van a ser menores que a nivel del mar.
Según se ha podido comprobar (observando los resultados de 67 estudios) que el
VO2MAX.
desciende en altitud de forma curvilínea (Fulco y cols. 1998) El efecto de la
altitud sobre el VO2MAX
se pueden empezar a notar a partir de los 500 metros de altura (sobre todo en
deportistas bien entrenados) pero es a partir de los 1.000-1.500 metros cuando
el VO2Max
disminuye un 1% cada 100 metros. Sin embargo, el efecto es más acusado a
grandes alturas lo que razona la relación curvilínea entre estos dos
parámetros.
No obstante, el VO2MAX
depende de la capacidad de suministro de oxígeno (factores centrales) y de la
capacidad de utilización del mismo (factores periféricos). Por esto, la
capacidad máxima de suministro de oxígeno (SumO2MAX)
viene determinada por el contenido de oxígeno en la sangre arterial (CaO2MAX)
durante el esfuerzo máximo y el gasto cardiaco máximo (QMAX). De esta manera
podemos decir que:
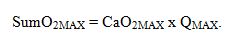
En hipoxia aguda el VO2MAX
está disminuido debido al descenso de la capacidad máxima del suministro de
oxígeno y se produce por diferentes causas dependiendo del grado de hipoxia. A
4.000 metros de altura o menos, la principal causa es el descenso de la
saturación de la hemoglobina con oxígeno, ya que el gasto cardiaco máximo es
el mismo que a nivel del mar. Sin embargo, a alturas superiores a los 4.000
metros, el descenso del VO2MAX
también se debe a un descenso del gasto cardiaco máximo. Y, por último, a
alturas superiores a 5.300 metros, el VO2MAX
es un 47% inferior al del nivel del mar y es causado por el descenso de la
saturación de oxígeno de la hemoglobina arterial (dos tercios) y al descenso
del gasto cardiaco máximo (un tercio) (Calbet y cols. (2003).
El descenso del VO2MAX
con la hipoxia es mayor en personas que disponen de un VO2MAX
más alto pero esto no excluye que el mismo siga siendo mayor que el de las
personas que lo tienen más bajo. No obstante las diferencias entre sujetos
entrenados y no entrenados disminuyen en altura.
Teniendo en cuenta el descenso del gasto cardiaco máximo, la capacidad máxima
de suministro de oxígeno mejora con la aclimatación hasta el punto que al
pasar una o dos semanas es la misma que la que se tenía a nivel del mar ya que
aumenta la concentración de hemoglobina y la saturación con oxígeno de la
misma. Como consecuencia de esto, el contenido de oxígeno de sangre arterial en
reposo es igual o superior en los sujetos aclimatados que los observados de
ellos mismos a nivel del mar.
En sujetos aclimatados durante nueve semanas a 5.260 metros la concentración de
hemoglobina aumentó un 34%, la saturación de la hemoglobina en el esfuerzo fue
de 72,5% y el gasto cardíaco fue un 15% inferior. Como consecuencia, se
aumentó el suministro máximo de oxígeno en un 54% al compararlos con los
valores observados en hipoxia aguda. No obstante el VO2MAX
sólo mejoró un 13% por lo que el que medimos en altitud fue un 26% inferior al
observado a nivel del mar (Calbet, 2006). Los estudios efectuados durante la
Expedición del Centro de Investigación Muscular de la Universidad de
Copenhague a Chacaltaya (Bolivia) en 1998 dieron repuesta a la pregunta de por
qué aumenta tan poco el VO2MAX
con la aclimatación a la altura si aumenta considerablemente el suministro
máximo. Estos estudios dieron como resultado que el aumento del suministro no
puede ser aprovechado por el músculo esquelético, ya que el flujo sanguíneo
muscular en hipoxia es muy inferior al observado en normoxia. Otra razón a la
que se le ha atribuido este suceso es que en hipoxia existe una limitación a la
difusión de oxígeno, debido a que el gradiente de presión parcial de oxígeno
entre los capilares musculares y la mitocondrias es menor pero está demostrado
que este no es el mecanismo responsable del poco aumento del VO2.
Otro dato importante sobre este segundo argumento es que en condiciones de
hipoxia se produce una alteración en la distribución del gasto cardiaco, es
decir, parte de la sangre se desvía a otros órganos que tienen menor necesidad
de oxígeno que los músculos esqueléticos.
7.
Efectos de la permanencia en altura sobre el VO2MAX
a la vuelta a nivel del mar
Si la estancia en altura no se realiza a altitudes superiores a los 2.500 metros
y durante tres semanas al menos, la masa eritrocitaria aumentara muy poco por lo
que el aumento de la concentración de hemoglobina se deberá mayoritariamente a
la hemoconcentración (disminución del volumen plasmático). Además, a las
48-72 horas del retorno a nivel del mar la concentración de hemoglobina se
acerca mucho a la observada antes de la exposición a altura debido a la
expansión del volumen plasmático. Como consecuencia, la capacidad máxima de
suministro de oxígeno es similar antes y después de la permanencia en altura
como demostró (Saltin, 1.996). Llegando a la conclusión de que la permanencia
en altura no se asocia a un aumento del VO2MAX
al regresar a nivel del mar.
8.
Efectos de la hipoxia sobre la fuerza muscular y la potencia muscular
Sólo puede haber una pérdida de fuerza y potencia muscular si se produce una
disminución de la masa muscular, cosa que ocurre con relativa frecuencia con la
permanencia prolongada a grandes altitudes (Ferretti y cols. 1990).
9.
Utilización de la hipoxia como medio para mejorar el rendimiento
9.1.
Entrenamiento en altura para mejorar la resistencia aeróbica
El principal argumento que defiende el entrenamiento en alturas moderadas para
mejorar la resistencia aeróbica es que se produce un aumento de la
concentración de hemoglobina. De esta manera, cuando se vuelva a nivel del mar
estará aumentada la capacidad de suministro de oxígeno y, junto a esto,
también aumentarán el VO2MAX
y la resistencia aeróbica. Sin embargo, este aumento de la concentración de la
hemoglobina se debe principalmente a una disminución del volumen plasmático
por lo que es un mecanismo muy eficaz para aumentar rápidamente el contenido de
oxígeno de la sangre. Una solución para evitar la deshidratación producida
por la disminución del volumen plasmático consiste en vivir en altura y
entrenar a nivel del mar. Según Robach y cols. (2002), después de una larga
exposición a hipoxia, la vuelta a nivel del mar acarrea una disminución de la
concentración de hemoglobina (a niveles similares o ligeramente superiores a
los del nivel del mar originales) que se debe a una expansión del volumen
plasmático que se produce por la retención de agua. A los 3-10 días tras la
vuelta a nivel del mar se vuelven a los valores anteriores a la exposición a la
hipoxia. Si la estancia en altura no supera el mes y la altura es inferior a los
3.000 metros los efectos sobre la concentración de hemoglobina no son
significativos y la vuelta a los valores normales tras la exposición a la
hipoxia se produce en tres días. Svendenhag y cols. (1997) estudiaron a
esquiadores de fondo a 1.900 metros de altura llegando a las siguientes
conclusiones: no se observaron cambios significativos en la masa total de
hemoglobina, a los once días después del regreso a nivel del mar el volumen
sanguíneo era un 7% mayor, la masa ventricular izquierda aumento un 10%. No se
sabe hasta qué punto influyo la hipoxia en estos resultados.
La mayoría de los estudios sugieren que el entrenamiento en altura no permite
aumentar el VO2
a nivel del mar pero sí el VO2
en altura posiblemente debido a la aclimatación. Los efectos sobre el
rendimiento del entrenamiento en altura no están muy claros aunque hay estudios
como el de Levine y Stray-Gundersen (1997) que demuestran una mejora de las
marcas después de entrenar en altura. Estos dos mismos autores alegaron que el
entrenamiento a nivel del mar es más efectivo que en altura porque nos permite
mantener la misma intensidad absoluta durante más tiempo y el tiempo de
recuperación entre series ha de ser mayor.
También corresponden a estos dos autores los estudios mejores diseñados para
analizar los efectos de distintas estrategias de entrenamiento en altura.
Descubrieron que la permanencia en altura y el entrenamiento en hipoxia moderada
son muy eficaces para aumentar el rendimiento en competiciones de 3.000 y 5.000
metros en deportistas de alto nivel y de élite. No ha sido posible discriminar
las mejoras que han tenido lugar por acción del entrenamiento y cuáles por la
acción de la hipoxia. También llama la atención que el incremento de la
concentración de la hemoglobina no correlacionó con el incremento del VO2MAX.
En general, un aumento de la concentración de hemoglobina se asocia a un
descenso de la concentración de lactato durante el esfuerzo submáximo.
También se han ensayado variantes del sistema HiLo (vivir en altura y entrenar
a nivel del mar) (Ashenden y cols. 1999) que consisten en estudiar el efecto de
dormir en hipoxia y entrenar a nivel del mar (a unos 600 metros de altura). En
este estudio no se observaron cambios significativos en la masa de hemoglobina.
9.2.
Adaptaciones musculares al entrenamiento en altura
En algunos estudios como los de Terrados y cols. (1990) y Melissa y cols. (1997)
se han observado aumentos más acusados de la actividad de las enzimas del
metabolismo oxidativo y de la mioglobina al entrenar en hipoxia y a la misma
intensidad absoluta. Pero esto solo es posible si la masa muscular sometida a
entrenamiento es pequeña porque por el contrario no se podría mantener la
misma intensidad absoluta en hipoxia que en normoxia.
También hay que tener en cuenta que en los sujetos entrenados la capacidad
oxidativa muscular es mayor a la capacidad de suministro de oxígeno, por lo que
si no aumento esta capacidad de suministro no tiene importancia que aumente la
capacidad oxidativa muscular.
Otra de las ventajas que le ha atribuido al entrenamiento en hipoxia son mejoras
en la mioglobina muscular y en la capilarización de las fibras musculares.
Estos no aumentan la capacidad de extracción de oxígeno cuando deberían
hacerlo. Esta capacidad depende de varios factores como la afinidad de la
hemoglobina por el oxígeno, el gradiente de la presión de oxígeno entre el
capilar y la mitocondria, posiblemente la concentración de mioglobina, el
tiempo medio de transición de la sangre a través de los capilares y la
densidad capilar. La afinidad de la hemoglobina por el oxígeno disminuye a al
aumentar la concentración eritrocitaria de 2-3 DPG con la hipoxia, lo que
debería facilitar la cesión de oxígeno al músculo si no se produjera
alcalosis respiratoria en hipoxia (desplaza la curva de disociación de la
hemoglobina hacia la izquierda) que dificulta la cesión de oxígeno en el
músculo, pero facilita la captación alveolar de oxígeno. El efecto de estos
dos sucesos deja la curva de disociación de la hemoglobina en una posición
similar a la observada a nivel del mar. Los efectos de la 2-3 BPG desaparecen
como mucho a los dos días del regreso al nivel del mar, mientras que la
densidad capilar expresada como número de capilares por fibra muscular no
aumenta con la sola permanencia en altura pero si lo hace o se mantiene
inalterada la densidad capilar expresada en número de capilares por mm2 de
sección muscular (debido a la atrofia de las fibras musculares) Lundby y cols.
2004).
9.3.
Efectos del entrenamiento en altura sobre la capacidad anaeróbica
El entrenamiento de alta intensidad en altura se asocia a un aumento de la
actividad enzimática relacionada con la glicólisis y la capacidad tampón.
Pero, por otro lado, la actividad de la bomba sodio-potasio y la reserva total
de bicarbonato disminuyen. Actualmente no se ha podido demostrar claramente una
superioridad del entrenamiento en altura o en condiciones de hipoxia para la
mejora de la capacidad anaeróbica, ya que hay demasiada controversia y errores
metodológicos en los estudios publicados sobre el tema.
9.4.
Hipoxia intermitente y rendimiento deportivo
El término intermitente se refiere a que los sujetos sólo son sometidos a
hipoxia durante una parte del día. Tras los estudios de Levine y
Stray-Gundersen apareció un gran interés por la estrategia de entrenamiento
HiLo (aunque no se considera una forma de hipoxia intermitente). Se han probado
dos principales modelos de hipoxia intermitente: modelos de hipoxia intermitente
continua y modelos de hipoxia intermitente discontinua. En el primero, los
sujetos pasan una parte del día viviendo en hipoxia (altitudes entre 2.200 y
3.000 metros o equivalente) y entrenan a nivel mar. Los pocos estudios
publicados sobre este tema presentan unos cambios hematológicos mínimos (no es
extraño porque se han sometido poco tiempo y a niveles de hipoxia bajos), pero
han presentado mejoras en la capacidad anaeróbica y mejoras en el rendimiento
medido como potencia media desarrollada en cuatro minutos (Roberts y cols.
2003).
La conclusión que podemos sacar de diversos artículos sobre la estrategia de
pasar la noche en alturas entre 2.500 y 3.000 metros y entrenar a nivel del mar
podría facilitar una mejora de las marcas entre un 0,8 y un 1% en competiciones
cuya duración se encuentre entre 45 segundos y cuatro minutos. Aunque esta
mejora parezca insuficiente no es tan irrelevante como parece, ya que se pueden
producir mejoras en más de un 20% en una carrera de 1.500 metros por ejemplo.
La hipoxia intermitente discontinua abarca a los procedimientos en los cuales se
somete a los sujetos a una o más sesiones diarias de una o dos horas de
duración durante la cual se repiten ciclos de hipoxia severa (equivalente a
altitudes entre 5.000 y 6.5000 metros) de corta duración (5 minutos)
intercalados con periodos también cortos (5 minutos) de normoxia. Este tipo de
entrenamiento no produce efectos sobre las variables hematológicas, el VO2MAX
o el rendimiento deportivo (Julian y cols. 2004).
Estudios como el de Adams y cols. (1975) o el de Ventura y cols. (2003) afirman
que el entrenamiento en hipoxia mientras se vive a nivel del mar no produce
mejora en el rendimiento superior a la conseguida a nivel del mar. Pero si es
cierto que el entrenamiento en hipoxia produce adaptaciones en enzimas
específicas cuya influencia en el rendimiento a nivel del mar no ha podido ser
demostrada aún.
9.5.
Cambios orgánicos de la permanencia en altura que pueden deteriorar el
rendimiento
La aclimatación a la altura puede ocasionar cambios que podrían resultar
perjudiciales para el rendimiento a nivel del mar. Por ejemplo, produce un
descenso del agua corporal total y del volumen plasmático que puede disminuir
el VO2MAX
y la resistencia aeróbica (sobre todo en ambientes cálidos) (Calbet, 2006).
Dormir a una altitud de superior a 3.000 metros y entrenar a nivel del mar
durante más tres semanas produce un descenso de la actividad de la bomba
sodio-potasio que podría afectar a la resistencia aeróbica y anaeróbica
(Calbet, 2006).
Con la exposición a la hipoxia, especialmente a alturas moderadas y elevadas,
se puede producir una pérdida importante de masa muscular, por lo que la
velocidad y la potencia muscular podrían verse mermadas también (Calbet,
2006).
La hipoxia severa también dificulta la actividad mental y las tareas motoras
finas pudiendo quedar efectos secundarios sobre la técnica (Calbet, 2006).
A altitudes superiores a 6.000 metros casi todos los sujetos presentan
alteraciones del sueño que pueden afectar al rendimiento deportivo (West y
cols. 1986).
10.
Conclusiones
Tras realizar esta revisión se pueden llegar a varias conclusiones:
-
Sin
que exista aclimatación el rendimiento en condiciones de hipoxia va a verse
disminuido. Por ejemplo, el VO2MAX
va a disminuir un 1% cada 100 metros.
-
El
rendimiento en las pruebas en las que se pongan en marcha únicamente
mecanismos anaeróbicos no van a verse afectadas por la hipoxia.
-
Va
a mejorar el rendimiento en pruebas en las que sea determinante el
rozamiento con el aire (saltos, lanzamientos,….) y en aquellas en las que
se consigan grandes velocidades (ciclismo de carretera).
-
El
entrenamiento en hipoxia es claramente eficaz para mejorar el rendimiento en
estas mismas condiciones, ya que se mejora notablemente el rendimiento en
pruebas de resistencia (hasta un 45%).
-
Con
la aclimatación a la hipoxia, la masa eritrocitaria puede aumentar hasta un
50% en ocho meses a partir de los 2.500-3.000 metros.
-
Con
la aclimatación se puede mejorar el VO2MAX
en un 13% en hipoxia pero no a nivel del mar.
-
Si
se entrena a menos de 2.500-3.000 metros, los eritrocitos se pierden en
24-48 horas.
-
En
hipoxia disminuyen las diferencias en el rendimiento entre sujetos
entrenados y no entrenados.
-
Permanencia
en hipoxia y el entrenamiento a nivel del mar o en hipoxia moderada es
eficaz para mejorar el rendimiento en competiciones de 3.000 y 5.000 metros
en deportistas de alto nivel o de élite.
-
Existen
cambios orgánicos perjudiciales derivados de la permanencia en altura que
pueden empeorar el rendimiento.
-
Hay
autores que recomiendan el entrenamiento a nivel del mar, ya que podemos
mantener una misma intensidad absoluta más tiempo que en altura.
-
De
todas formas, los efectos sobre el rendimiento del entrenamiento en altura
no están muy claros actualmente.
En resumen, el entrenamiento en altura es claramente eficaz para aumentar el
rendimiento deportivo cuando la competición va a tener lugar en altura. Para
mejorar en rendimiento a nivel del mar, la combinación de permanencia en altura
con entrenamiento de alta intensidad a 1.250 metros y entrenamiento de moderada
y baja intensidad entre 2.000 y 3.000 metros ha originado mejores resultados en
algunos estudios. Pero todavía se necesitan más investigaciones para
determinar si el entrenamiento en altura puede ofrecer alguna ventaja para
competir a nivel del mar en disciplinas de resistencia aeróbica.
Bibliografía
-
Adams,
W.C., Bernauer E.M., Dill D.B., Bomar, J.B Jr . (1975). Effects of
equivalent sea-level and altitude training on VO2max
and running performance. J Appl Physiol (39), 262-6.
-
Ashenden,
M.J. Gore, C.J., Martin, D.T., Dobson, G.P, Hahn A.G. (1999). Effects of a
12-day "live high, train low" camp on reticulocyte production and
hemoglobin mass in elite female road cyclists. Eur J Appl Physiol Occup
Physiol (80), 472-8.
-
Boushel,
R., Calbet, J.A., Radegran, G., Sondergaard, H., Wagner, P.D., Saltin,
B.(2001). Parasympathetic neural activity accounts for the lowering of
exercise heart rate at high altitude. Circulation (104), 1785-91.
-
Calbet,
J.A., Bouschel, R., Radegran, G., Sondergaard, H., Wagner, P.D., Saltin,
B.(2003b). Why is VO2MAX
after altitude acclimatization still reduced despite normalization of
arterial O2
content?. Am J Physiol Regul Integr Comp Physiol. (284), 304-16.
-
Calbet,
J.A., Boushel, R., Radegran, G., Sondergaard, H., Wagner, P.D., Saltin,
B.(2003a). Determinants of maximal oxygen uptake in severe acute hypoxia. Am
J Physiol Regul Integr Comp Physiol (284), 291-303.
-
Calbet,
J.A., Radegan, G., Bouschel, R., Sondergaard, H., Saltin, B., Wagner,
P.D.(2004). Plasma volume expansion does not increase maximal cardiac output
or VO2MAX
in lowlanders acclimatized to altitude. Am J Physiol Heart Circ Physiol
(287), 1214-24.
-
Calbet,
J.A. (2006). Fisiología de la altitud y ejercicio físico. En: López
Chicharro, J., Fernández Vaquero, A., Fisiología Del Ejercicio 3ª ed.
(696-637). Buenos aires; Madrid: Médica Panamericana.
-
Eckardt,
K.U., Boutellier, U., Kurtz, A., Schopen, M., Koller, E.A., Bauer, C.
(1989). Rate of erythropoietin formation in humans in response to acute
hypobaric hypoxia. J Appl Physiol (66), 1785-8.
-
Dempsey,
J.A., Foster, H.V.(1982) Mediation of Ventilatory Adaptations. Physiol
Rev (62), 262-346.
-
Ferretti,
G., Hauser, H., Prampero, P.E. (1990). Maximal muscular power before and
after exposure to chronic hypoxia. Int J Sports Med (11 Suppl 1),
31-4.
-
Fulco,
C.S., Rock, P.B., Cymerman, A. (1998). Maximal and submaximal exercise
performance at altitude. Aviat Space Environ Med (69), 793-801.
-
Ge,
R.L., Witkowski, S., Zhang, Y., Alfrey, C., Sivieri, M., Karlsen, T. y cols.
(2002). Determinants of erythropoietin release in response to short-term
hypobaric hypoxia. J Appl Physiol (92), 2361-7.
-
Ghofrani,
H.A., Reichenberger, F., Kohstall, M.G., Mrosek, E.H., Seeger, T.,
Olschewski, H. y cols. (2004). Sildenafil increased exercise capacity during
hypoxia at low altitudes and at mount Everest base camp: a randomized,
double-blind, placebo controlled crossover trial. Ann Intern Med
(141), 169-77.
-
Hannon,
J.P., Shields, J.L., Harris, C.W. (1969). Effects of altitude
acclimatization on blood composition of women. J Neurophysiol (26),
540-7.
-
Julian,
C.G., Gore, C.J., Wilber, R.L., Daniels, J.T., Fredericson, M.,
Stray-Gundersen, J. y cols. (2004). Intermittent normobaric hypoxia does not
alter performance or erythropoietic markers in highly trained distance
runners. J Appl Physiol (96), 1800-7.
-
Lawler,
J., Powers, S.K., Thompson, D. (1988). Linear relationship between VO2MAX
decrement during exposure to acute hypoxia. J Appl Physiol (64),
1486-92.
-
Levine,
B.D., Stray-Gundersen, J. (1997). "Living high-training low":
effect of moderate-altitude acclimatization with low-altitude training on
performance. J Appl Physiol (83), 102-12.
-
Lundby,
C., Araoz, M., Van Hall, G. (2001) Peak heart rate decreases with increasing
severity of acute hypoxia. High Alt Med Biol (2), 369-76.
-
Lundby,
C., Calbet, J.A., Van Hall, G., Saltin, B., Sander, M. (2004). Pulmonary gas
exchange at maximal exercise in Danish lowlanders during 8 wk of
acclimatization to 4,100 metros and in high-altitude Aymara natives. Am J
Physiol Regul Integr Comp Physiol (287), R1202-8.
-
Lundby,
C., Van Hall, G. (2004). Lactate metabolism at high altitude. High Alt
Med Biol (Summer; 5),195-6; author reply 197-8.
-
Melissa,
L., MacDougall, J.D., Tarnopolsky, M.A., Cipriano, N., Green, H.J. (1997).
Skeletal muscle adaptations to training under normobaric hypoxic versus
normoxic conditions. Med Sci Sports Exerc (29), 238-43.
-
Milledge,
J.S., Cotes, P.M. (1985). Serum erythropoietin in humans at high altitude
and its relation to plasma renin. J Appl Physiol (59), 360-4.
-
Reynafarje,
C., Lozano, R., Valdivieso, J. (1959). The polycythemia of high altitude:
iron metabolism and related aspects. Blood (14), 433-55.
-
Richalet,
J.P., Gratadour, P., Robach, P., Pham, I., Dechaux, M., Joncquiert-Latarjet,
A. y cols. (2004). Sildenafil inhibits the altitude-induced hypoxemia and
pulmonary hypertension. Am J Respir Crit Care Med.Robach, P., Lafforgue, E.,
Olsen, N.V., Dechaux, M., Fouqueray, B., Westerterp-Plantenga, M. y cols.
(2002). Recovery of plasma volume after 1 week of exposure at 4,350 m.
Pflugers Arch (444), 821-8. Epub 2002 Aug 2.
-
Roberts,
A.D., Clark, S.A., Townsend, N.E., Anderson, M.E., Gore, C.J., Hahn, A.G.
(2003). Changes in performance, maximal oxygen uptake and maximal
accumulated oxygen deficit after 5, 10 and 15 days of live high: train low
altitude exposure. Eur J Appl Physiol (88), 390-5. Epub 2002 Nov 7.
-
Saltin,
B. (1996). Exercise and environments: focus on altitude. Res Q Exerc
Sport (67), S1-10.
-
Sánchez,
C., Merino, C., Figallo, M. (1970). Simultaneus measurements of plasma
volume and cell mass in polycythemia of high altitude. J Appl Physiol
(28), 775-8.
-
Svedenhag,
J., Piehl-Aulin, K., Skog, C., Saltin, B. (1997). Increased left ventricular
muscle mass after long-term altitude training in athletes. Acta Physiol
Scand (161), 63-70.
-
Terrados,
N., Jansson, E., Sylven, C., Kaijser, L. (1990). Is hypoxia a stimulus for
synthesis of oxidative enzymes and myoglobin? J Appl Physiol (68),
2369-72.
-
Ventura,
N., Hoppeler, H., Seiler, R., Binggeli, A., Mullis, P., Vogt, M. (2003). The
response of trained athletes to six weeks of endurance training in hypoxia
or normoxia. Int J Sports Med (24), 166-72.
-
West,
J.B., Peters, R.M. Jr., Aksness, G., Maret, K.H., Milledge, J.S., Schoene,
R.B. (1986). Nocturnal periodic breathing at altitudes of 6,300 and 8,050 m.
J Appl Physiol (61), 280-287.
-
Weil,
J.V., Jamieson, G., Brown, D.W., Grover, R.F. (1968). The red cell
mass-arterial oxygen relationship in normal man. J Clin Invest (47),1627-39.
-
Zaccaria,
M., Rocco, S., Noventa, D., Varnier, M., Opocher, G. (1998) Sodium
regulating hormones at high altitude: basal and post-exercise levels. J
Clin Endocrinol Metab (83), 570
Otros artículos sobre Actividad
Física y Salud
 |
|
|
EFDeportes.com, Revista
Digital · Año 16 · N° 161 | Buenos Aires,
Octubre de 2011
© 1997-2011 Derechos reservados
|
