Los humanos, salvo raras excepciones, pasamos toda nuestra vida en sociedad,
encontrando en las señales verbales y no verbales, códigos de comunicación
intraespecífica que nos ha permitido trasladar fácilmente la información
disponible de unos a otros. Estas interrelaciones incluyen conductas que
expresan nuestra posición social (percepción del estatus por otros miembros),
y que parece ser producto de una memoria social, que Dubar (2003)
definiría como la capacidad de recordar encuentros y percibir rangos de
jerarquía.
En primates, las diferencias intragrupo en los niveles de cortisol, son
estadísticamente significativos en los subordinados, más propensos a la
hipercortisolemia que los dominantes (Abbot et al, 2003).
Para Sapolsky (2004), estas respuestas endocrinas de los subordinados suelen
ocurrir, pero no siempre, ni de manera universal en todas las especies. La
relación entre el bienestar y el estatus social, puede verse afectada
drásticamente por las estrategias de afrontamiento individual, tanto para
humanos como para animales. Por tanto, las diferencias sociales y culturales (en
el caso del hombre) pueden tener origen y consecuencia en el estrés
psicosocial, lo cual dificulta una respuesta estandarizada en base a conceptos
exclusivamente psicofisiológicos, como apuntaba Seyle (Seyle, 1950). Sin
embargo, existen condiciones sociales que pueden actuar como agentes estresantes
y que modifican el estatus social y la salud (Sandín, 2003).
En comunidades humanas, y en otras especies gregarias jerarquizadas, es
inevitable convivir para sobrevivir, y también es inevitable convivir sin
conflictos de intereses. La forma en que éstos se resuelven suele ser mediante
la competición por la obtención del recurso. El análisis, en el mundo animal,
de las interrelaciones, mediaciones en conflictos, competencia, estructura
social, etc., ha permitido conocer conceptos que ayudan a encontrar respuestas
en los mecanismos sociales, psicológicos (percepción, evaluación,
afrontamiento y personalidad) y, en los últimos años, neuroendocrinos; y
algunos síndromes conductuales. Es interesante que la investigación sobre la
conducta humana, siga una línea similar, con el objetivo de analizar la
complejidad de los comportamientos sociales (Sapolsky, 2004).
La necesidad de estudiar la motivación, los procesos cognitivos, las variables
psicológicas, la atribución causal de los resultados o la satisfacción, ha
podido explicar algo la enorme varianza que se obtiene de los resultados de las
respuestas neuroendocrinas en la competición humana (Salvador, 2005; Salvador y
Costa, 2009; Archer, 2006; Filaire et al, 2009; Van Anders y Watson, 2006,
2007).
El eje
HPG desde un punto de vista ecológico y evolutivo
Altos niveles de T y dominancia social han sido la base de muchos estudios
científicos. La relación entre la T y algunas conductas sociales o rangos de
dominancia intraespecífica, se ha confirmado en pájaros (Buchanan et al,
2010), primates (Abbot et al, 2003; Mehta et al, 2008; Sapolsky, 1991) y
también en seres humanos (Sapolsky, 2004; Gillespie, 1971; Wingfield, Hegner,
Duffy y Ball, 1990, Archer, 2006; Marzur y Lamb, 1980).
Cuando los seres humanos con niveles altos de T, ganan o pierden en competición
deportiva o social, la derrota aumenta los niveles de C y la victoria los
disminuye, generalmente. Para los participantes con bajos niveles de T, la
victoria o el fracaso no aporta cambios significativos en los niveles de C,
aunque sean ligeramente descendentes en los ganadores (Salvador y Costa, 2009;
Salvador 2003; Salvador, 2005). Para Blanchard (2001), cuando se obtiene la
victoria, las probabilidades de éxito en las competiciones posteriores
aumentan. La victoria otorga una posición de liderazgo jerárquico sobre los
perdedores, incluidos cambios de carácter fisiológico, conductual y de
dominancia sobre los otros.
La hipótesis de estatus biosocial (BSSH) en primates de Marzur y Lamb
(1985), insta a pensar que la victoria de un enfrentamiento produce aumentos
significativos en los niveles de T, mientras que la derrota disminuciones;
además el interés del ganador en participar en nueva conductas agonísticas se
incrementaban (Figura 1). Determinaron que los cambios en la T
continuaban siendo diferentes en base a la consecución de la meta (Marzur y
Lamb, 1980). Esta hipótesis ha sido estudiada en deportes de raqueta, donde los
tenistas perdedores durante la competición deportiva, tienen mayores niveles de
C basal y durante el torneo que los ganadores (Filaire et al, 2009).
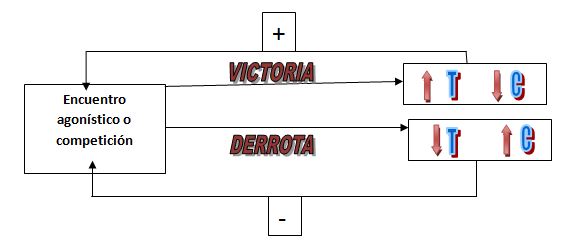
Figura
1. Representación de la hipótesis del estatus biosocial de Marzur,
basado en la relación recíproca
entre
hormonas y comportamiento. (Adaptado de Marzur y Lamb, 1985 y Marzur y Booth,
1998)
La HC recogida por muchos estudios científicos en el ámbito del deporte,
sería la tendencia general de los organismos implicados en luchas por el
recurso, quienes ven aumentados sus niveles de T antes de la contienda
intraespecífica. Los animales ganadores de estos enfrentamientos, reciben la
retroalimentación propia de la T, como el cambio en los fenotipos conductuales
y físicos, mejora de la autoestima y percepción positiva de poder alcanzar una
nueva meta; entre éstos, los procesos anabolizantes de la T mejoran la
mineralización ósea y el aumento de la masa muscular (Nowak et al, 2010).
El eje
HPA desde un punto de vista ecológico y evolutivo
En humanos, el eje HPA puede ser activado por gran cantidad de estresores, los
de tipo psicológico son los más comunes, en oposición a los fisiológicos o
sistémicos. Muchas de las respuestas disponibles son de carácter
anticipatorio, o sea, que están basadas en las expectativas de resultados que
poseen en la memoria vital, y juegan un importante rol en los modelos de estrés
crónico, afectando al cerebro, la conducta y los procesos cognitivos (Lupien et
al, 2009, Sapolsky, 1992).
Figura
2. Circuitos responsables de la respuesta
neuroendocrina de estrés
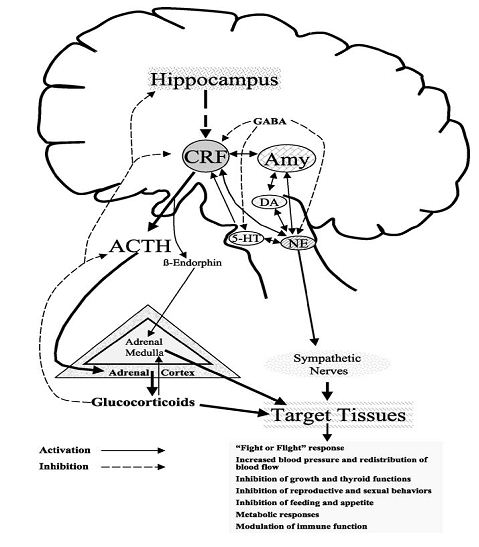
FCI
= Factor de liberación de Corticotropina (Núcleo Hipotalámico
Paraventricular);
5-HT
=Serotonina (Núcleo Dorsal); NE
= Noradrenalina; Amy= Amígdala; DA=Dopamina (Sistema
Mesolímbico)
(Adaptado
de Carrasco y Van de Kar, 2003)
Los componentes neuroendocrinos activados por los estresores incluyen el
incremento de los niveles de adrenalina y noradrenalina desde el sistema
nervioso simpático y la médula adrenal. La CRF y vasopresina desde la zona
paraventricular y, unos segundos después, la pituitaria secreta
adenocorticotropina (ACTH), fomentando el aumento de glucocorticoides desde la
glándula adrenal (Figura 2.)
Costes
de la Carga Alostática
La Carga Alostática se refiere al coste acumulativo para el cuerpo, de los
ajustes individuales, en base a su morfología, fisiología y conducta ante los
cambios ambientales, estado social, etc.; puede verse variado a lo largo de las
diferentes etapas del ciclo vital de cada animal.
Cuando se enfría el cuerpo éste produce calor, cuando está amenazado por
bacterias se producen anticuerpos, y cuando existe un peligro externo el
organismo se prepara para llevar a cabo una respuesta de lucha o huída
(Sandín, 2003).
La sobrecarga alostática puede ser: un estado en el cual los costes de cada
fase del ciclo vital y las situaciones concomitantes exceden los recursos
disponibles para proveer al sistema de energía suficiente (tipo I); o un estado
en el que ponemos a prueba nuestro cuerpo con estados nocivos que pueden llegar
a ser crónicos (Tipo II); este último tipo, es el que se produce en los seres
humanos, y puede llegar a provocar patologías graves si no se alivia la carga
de algún modo (Mc Ewen y Wingfield, 2003; Bower et al, 2008).
En el tipo II, la sobrecarga alostática es demasiado alta como para que el
consumo de energía sea suficiente para retomar el equilibrio fisiológico. Como
consecuencia de ello, la secreción de Glucocorticoides es crónica y elevada y,
junto con los desequilibrios de otros mediadores, con el paso de los años
desencadenan alguna patología. Este tipo de sobrecarga es la más común en el
ser humano y en los animales en cautiverio. No favorece respuesta de
escape-huida; salir de ella sólo es posible cuando, los seres humanos que
sufren conflictos sociales, aprenden a cambiar su propio comportamiento y,
sujetos a las exigencias del orden establecido, alivian las condiciones sociales
negativas con un estilo de afrontamiento positivo de los eventos vitales
(Wingfield, 2005, Sapolsky, 2004, Moya-Albiol et al, 2005, Roy et al, 2003;
Thompson, Mata, Jaeggi, Buschkuehl y Jonides, 2010).
La carga alostática, como veremos también en el ámbito del deporte, repercute
directamente sobre el metabolismo energético, el sistema cardiovascular, el
sistema inmunológico, los estados de ánimo, etc. (Salvador y Costa; 2009;
Salvador et al, 2003; Salvador et al, 1995; Alix-Sy, Le Scanff y Filaire, 2008;
Ruiz et al, 2004; González-Bono, 1999; Filaire et al, 2009). Pero el estrés
desde el punto de vista darwiniano, no sólo debe entenderse como producto del
coste alostático; también, como un beneficio para la propia adaptación a un
entorno en continuo cambio, a través de la alostasis (Wingfield, 2005; Bower et
al, 2008).
La
respuesta neuroendocrina en competición deportiva
Alicia Salvador, (en su revisión de 2009) sobre el afrontamiento de la
competición en humanos, tras analizar los diferentes estudios iniciales sobre
la respuesta neuroendocrina, llegó a la conclusión de que existen una serie de
pautas que se repiten significativamente en la competición humana,
desarrollando el siguiente modelo hipotético de respuesta psicofisiológica en
deportistas (figura 3).
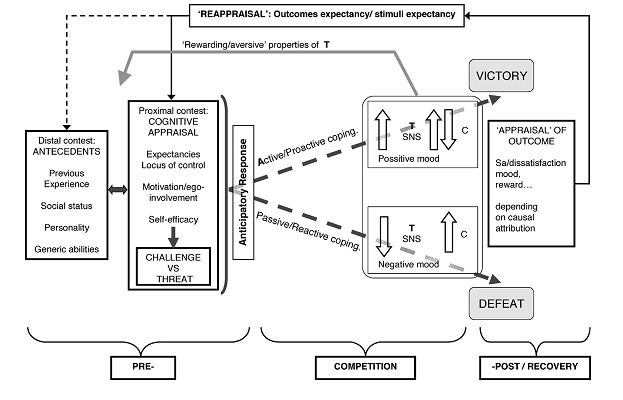
Figura
3. Modelo de respuesta neuroendocrina a la competición (Salvador y Costa,
2009)
En el modelo propuesto por Salvador y Costa (2009),
diferencia tres partes fundamentales de la competición deportiva en base a la
respuesta psicofisiológica:
-
La
fase precompetitiva, marcada por los antecedentes y su relación con la
situación propia de la competición; o lo que es lo mismo, diferencia entre
lo que la variable debe ser y lo que es realmente; como indica la Teoría de
la Activación Cognitiva del estrés (CAST), que provoca una respuesta
hormonal anticipatoria.
-
La
fase competitiva o de interrelación, donde el afrontamiento
activo o pasivo de la situación juega un papel determinante en el resultado
final.
-
La
fase postcompetitiva, marcada por una respuesta hormonal determinada
principalmente por el resultado final del encuentro agonístico y la
importancia que el deportista le da al resultado o en base a su locus de
control. La retroalimentación de la T y el C, mejorará las capacidades
psicofisiológicas del deportista para afrontar nuevos retos, o en su
defecto las disminuirá y afectará a los estados de ánimo (Salvador,
2009).
En concordancia con la CAST, sendos estudios han demostrado que la activación
de los ejes HPG y HPA, tienen una vinculación necesaria y determinante en seres
humanos sujetos a conductas competitivas. Carré desarrolló un estudio sobre
una muestra de jugadores de Hockey profesionales, a los que se les presentaba un
vídeo de un partido, jugado y ganado por el mismo equipo, previo al encuentro a
disputar; observándose un aumento significativo de la T precompetitiva (entre
un 42 y un 44 %) en los sujetos experimentales. Los procesos cognitivos, por
tanto, juegan su papel en la preparación del deportista para afrontar el
encuentro (Carré, Gilchrist, Morrisey y McCormick, 2010).
Importante es destacar, que los jugadores experimentados, suelen regular su
esfuerzo a lo largo de la temporada, con el fin de mantener el estado óptimo
competitivo, sin que los niveles de C o T puedan suponer una carga alostática
mantenida e incesante; de otro modo, y como hemos visto, los costes superarían
a los beneficios y con ello llegaría el sobreentrenamiento (Urhausen, Gabriel y
Kendarmanny, 1995) o los estados de ánimo negativos (Perna et al, 1998;
Moya-Albiol, Salvador, Costa, Ferrand y Lac, 2001).
Filaire et al (2001), también sobre una muestra de judokas, concluyó que las
relaciones existentes entre hormonas y variables psicológicas, dependían de la
respuesta, incidiendo en la importancia de analizar las estrategias de
afrontamiento, aspectos psicológicos previos y estados de ánimo.
En otros deportes individuales, en este caso de raqueta, se apreciaban aumentos
de la T precompetitiva, en relación con actividades deportivas no competitivas
(Edwards y Kurlander, 2010). Parece que la respuesta psicofisiológica es más
acusada en los deportes individuales, existiendo sendos estudios con deportes de
raqueta como el tenis (Booth, Shelley, Marzur, Tharp y Kittok, 1989).
Testosterona
y competición
Los deportistas expertos, que son exactamente la muestra que más interesada
podría estar en el estudio de sus respuestas hormonales a la competición,
tienen una mayor capacidad para mantener los niveles psicofisiológicos en
equilibrio, con el objetivo de finalizar la tarea o alcanzar un estadio óptimo
durante la temporada. Así, sobre una muestra de jugadores de baloncesto de
élite, Martínez y colaboradores, estudiaron los cambios hormonales en un
equipo profesional en la temporada competitiva. Los resultados confirman la idea
de una autorregulación del deportista experimentado, que no arrojaron
diferencias significativas en los niveles de C y T en los diferentes meses de
competición (Martínez et al, 2010).
Sin embargo, se han hallado resultados que instan a pensar, que los deportistas
afrontan con diferencia las competiciones por equipos y las individuales; siendo
las segundas más susceptibles de respuestas endocrinas más marcadas (Oxford,
Ponzi, Geary, 2009; Wagner, Flinn y England, 2009)
En laboratorio, se evaluaron los niveles de T y C en una competición masculina
no deportiva, indicando los resultados que la percepción de ganar o perder, y
la seguridad en la finalización de la tarea, se correlacionaba con la T, pero
no con el C; más cercano a cambios en los estados de ánimo o estatus social
(Glaude, 1989). Sin embargo, cuando la tarea implicaba el trabajo en equipo, los
individuos más relevantes en la victoria, generaban más T que los que
aportaron menos. Los individuos con altos niveles de T, generaban más C cuando
la competición se desarrollaba intragupo (Oxford et al, 2009).
La
Hipótesis del Aspirante (CH)
Originalmente propuesta para asociar la agresión y la T en pájaros monógamos
(Wingfield et al, 1990); se demostró que los niveles de T aumentaban en la
época de cría y durante los desafíos de otros machos en dicho contexto
(Winfield et al, 2001), en mamíferos (Wingfield, et al, 1990), donde se han
detectado cambios en el comportamiento y hormonales; y en primates (Muller y
Wranghan, 2004). En la investigación de Muller y Wranghan (2004), el encuentro
con hembras fértiles y receptivas, por parte de los machos, el aumento de la
agresividad se correlacionaban con la T. En los dominantes, estos niveles de T
son más altos que en los subordinados, los estatus sociales y los fenotipos de
fuerza y vitalidad, son consecuencia de ello; y por último, también en
humanos, sin encontrar diferencias significativas entre géneros (Archer, 2004,
Salvador y Costa, 2005).
Existe una considerable variación entre humanos y animales, ya que la agresión
queda inhibida a favor de otros conceptos selectivos, como por ejemplo las
preferencias del sexo opuesto, o los procesos cognitivos… por tanto, en
humanos debemos ser prudentes, e incorporar las diferencias individuales al
analizar la CH (Archer y Mehdikhani, 2003).
Un estudio interesante en competición controlada en laboratorio, a través de
una tarea con ordenador, en una muestra de 84 personas, analizó la respuesta
hormonal de los jugadores según el estado emocional de sus oponentes. Los
resultados instaban a pensar que la competición, independientemente del
resultado, aumentaba los niveles de T en ambos participantes, pero aumentaban
significativamente cuando los niveles de autoconfianza del oponente eran altos
(Van der Meij et al, 2010). Por el contrario, si la percepción del oponente,
sobre la importancia de la competición era baja, la T de los perdedores
disminuía en concordancia con la CH.
En humanos, rechazar o mantener la HC, depende de precursores evolutivos, pero
nuestros procesos cognitivos intervienen en las respuestas hormonales, como
hemos visto en la CAST (Salvador et al, 2009). Por tanto, cuestiones como la
percepción subjetiva de superioridad o el prestigio social, son directa e
inversamente proporcionales, respectivamente, a las conductas agresivas (Johnson
et al, 2007).
La posible confirmación de la HC, se aportaría analizando las respuestas
neuroendocrinas en las interacciones sexuales y competitivas (Marzur y Lamb,
2004; Salvador et al, 2003; González-Bono et al, 1999, 2000). La base de la
hipótesis es encontrar la proporción costes vs beneficios: las consecuencias
fisiológicas y conductuales de altos niveles de T mantenidos durante cierto
espacio de tiempo, y los beneficios que aporta para la supervivencia,
procreación, estatus social o conductas agonísticas (Wingfield et al, 2001).
Aunque inicialmente, la CH fue estudiada en el género masculino de las
diferentes especies analizadas, existe algún estudio sobre el impacto
psicofisiológico de la competición en el género femenino (Bateup et al, 2002;
Archer, 2004, 2005); donde los niveles de T aumentaban significativamente igual
que en el género masculino, cuando las féminas se enfrentaban una
competición.
La
competición deportiva
Salvador y colaboradores (2003) estudiaron la respuesta a la T en jóvenes
judokas, sin encontrar diferencias estadísticamente significativas entre
ganadores y perdedores de los encuentros. Este estudio, que fue realizado en
competición real, las diferencias hormonales de los participantes, y la
imposibilidad de replicar conductas y situaciones competitivas similares para
todos en cada combate, fue determinante para la obtención de resultados sin
diferencias sensibles.
Suay et al (1999), sobre una muestra de 26 judokas, estudiaron la respuesta
psicofisiológica a la victoria o la derrota, mostrando los ganadores alta
motivación a la tarea y satisfacción con los resultados, sin diferencias
significativas en los niveles de T. Similares resultados encontramos en Salvador
et al (1995); donde la actividad física aumenta los niveles de T y C, pero la T
aparece inversamente relacionada con indicadores de malestar psicológico:
ansiedad y estados de ánimo negativos.
Serrano et al (2000), en un estudio realizado sobre doce judokas en competición
oficial, confirmaron que la T se relacionaba con la autosatisfacción con la
tarea y la atribución del resultado al esfuerzo personal. Filaire et al (2001),
estudiaron también en doce judokas en competición oficial, pero en este caso
los niveles de T no variaban de manera significativa de unas competiciones a
otras, siendo el C quien correlacionaba positivamente con CA, SA y negativamente
con SC. En un estudio con futbolistas de diferentes niveles, también
concluyeron en la idea de que la respuesta emocional en competición estaba
relacionada con los niveles de C (Alix-Sy et al, 2008).
González-Bono et al (1999) estudiaron a 21 participantes, jugadores de
baloncesto de la Liga Nacional EBA, concluyendo que en los deportes de equipo,
en competición real, no se aprecian diferencias en los niveles de T, como
establecería la BSSH de Marzur; pero sí se apreció una afectación
negativa de los estados de ánimo en los perdedores.
Testosterona
y género femenino
Hay evidencias de que el género femenino responde con incrementos de
testosterona en situaciones de competición, en el mismo sentido que el género
masculino; lo que sugiere que las diferencias son individuales y más
influenciadas por el éxito o por las estrategias de afrontamiento activo o
pasivo, que por el género (Archer et al, 2005). Existe respuesta hormonal a los
desafíos en el este género, en concordancia con las BSSH y CH, con aumentos
significativos de la T precompetitiva (Hameishi al, 2007) y relacionado a la
victoria o la derrota (Oliveira, Gouveira, Oliveira, 2009). Como hemos
comentado, los incrementos o disminuciones de los niveles de T están más
influenciados por las diferencias individuales o la afiliación social, que con
el sexo (Kiulighan et al, 2004). En un estudio sobre una muestra de deportistas
femeninas en la modalidad de adiestramiento canino de Agility, se comprobó que
no manifestaban niveles significativos de C entre W y L (Mehta et al, 2008); en
este estudio se desarrolló una segunda investigación en laboratorio, donde el
objetivo teórico de la tarea con ordenador era demostrar el coeficiente
intelectual de cada una, en este caso, el género femenino se tomó la tarea
más en serio que la investigación anterior, ofreciendo diferencias
significativas intersujetos con altos niveles de T y aumentos o disminuciones
del C relacionados a la victoria o la derrota.
Testosterona
y estatus social (dominancia vs prestigio)
Carré et al (2001), estudiaron la relación entre la agresión y la conducta
competitiva en una muestra de participantes en una tarea con ordenador,
observando que la conducta agresiva está continua y directamente relacionada
con la recompensa que proporciona la T, regulando el interés por nuevas
conductas agonísticas, en concordancia con la BSSH de Mazur.
Las recientes tendencias estudian la diferencias individuales en la
sensibilización de los receptores cerebrales, su relación con los niveles de T
y la influencia transversal en el comportamiento (Manning, Bundred, Newton y
Flanagan, 2003); los estudios neurogenéticos especifican los posibles
coeficientes de heredabilidad de los fenotipos de conducta social, lo cual
refuerza la idea de una sensibilización individualizada en humanos (Craig y
Halton, 2009; Ebstein et al, 2010).
Otro aspecto muy interesante en las relaciones que se establecen entre estatus
social y toma de decisiones, es el desarrollado por Carney y Manson (2010),
quienes analizaron la relación entre la T y el papel que juega, junto a los
conceptos morales, en las decisiones que se deben tomar y que implican al bien
común frente al bien individual. Los individuos con altos niveles de T parece
dispuestos a respaldar y asumir decisiones morales que afecten a terceros.
Sugieren que estos participantes poseen una relación inversa entre T y
comportamiento empático; esto se estudió anteriormente en ratones de
laboratorio por Hermans et al (2006), mediante administración exógena de T. En
el género femenino, existen muchos trabajos que indican que los dos géneros
responden a la competitividad y la agresión con aumentos significativos de T
(Cashdan, 2003; Edwards y Kurlander, 2010); como resultado de la respuesta
hormonal a situaciones que implican a los recursos, estatus social, reproductivo…
Estas conductas tiene una relación cierta con procesos de carácter biológico,
y se ven afectados por los procesos cognitivos, emocionales y fisiológicos
(McAndrew, 2009).
En el ámbito social humano, estos trabajos nos instan a pensar porqué las
personas con altos niveles de T basal triunfan en el entorno socioeconómico y
en contextos que impliquen luchas por el control del estatus social. Esto es de
suma importancia para comprender, según Carney y Manson (2010), los mecanismos
que relacionan a triunfadores de Wall Street y trabajadores estresados;
políticos de diferentes ideologías y capacidad de afrontar situaciones
críticas; o a deportistas que ocupan los primeros puestos del ranking y su
control sobre esa posición preferencial en el entorno social deportivo.
En este sentido, los estudios de Johnson, Burk y Kirkpatrick (2007), distinguen
dos vías para la superioridad de unos humanos sobre los otros: la dominación
y el prestigio, el término prestigio ya fue acuñado por el
propio autor (Kirkpatrick, Waugh, Valencia, Webster, 2002). Hemos visto como,
indistintamente del género, la dominancia jerárquica se relaciona
positivamente con la T y ésta con la agresión; sin embargo, el prestigio se
correlaciona inversamente con la T, lo que indicaría cierta inhibición del eje
HPG en la superioridad jerárquica por esta vía. Esta variabilidad de la
conducta competitiva, no agresiva, basada en las metas de logros, podría ser
una vía adaptativa interesante para su estudio y análisis (Johnson et al,
2007). Mehta et al (2009), explican que los resultados son desiguales cuando se
trata de analizar si la T es facilitadora de las metas deportivas, indicando que
los niveles de T son directa e inversamente proporcionales si la competición es
individual o por equipos, respectivamente. La idea de una respuesta
psicofisiológica es mayor entre deportistas de alta competición individual,
con respecto a la competición por equipos, podría estar relacionado con
conceptos evolutivos de base, ya que la cooperación implica un importante grado
de inhibición de la agresividad intragrupo.
El eje
HPA y la competición
Las investigaciones con ratones no agresivos, ha mostrado mayores niveles de
cortisolemia y pérdida de peso, que en aquellos con conductas agresivas, tras
la exposición a oponentes agresivos. Estas manifestaciones producen
modificaciones del fenotipo en pájaros, donde la coloración y ornamentación
son consecuencia última de la capacidad de adaptación a los cambios y del
acceso preferencial a los recursos (Bartolotti et al, 2009); como en primates,
donde el estatus social está relacionado con los niveles de C, superiores en
los subordinados, quienes reciben menos atenciones, contacto y colaboración por
parte del grupo (Abbot et al, 2003); finalizando en los humanos, donde el
estrés ha sido directamente relacionado con patologías físicas y psíquicas,
y donde la cortisolemia queda relacionada a estatus socioeconómicos deprimidos
(Sapolsky, 2004).
La activación laboral, o las exigencias del entorno, son motivos para los
aumentos de C. Moya-Albiol el al (2005) compararon la respuesta hormonal,
emocional y cardiovascular de un grupo de profesores durante la jornada laboral
y los días de descanso. Durante el desempeño de sus trabajos, la percepción
de estrés y la frecuencia cardíaca aumentaban, deteriorándose al tiempo los
estados de ánimo positivos.
Se deduce, que el estrés juega un papel determinante en los reajustes
adaptativos del organismo ante los eventos vitales (situación laboral, social,
o en la alta competición deportiva, donde a veces se reúnen características
de todas las anteriores: mejora la calidad de vida, el reconocimiento social, y
en el caso de los deportistas profesionales, el estatus socioeconómico), y que
podría existir una predisposición epigenética de sensibilización al estrés.
Si así fuera, ello podría ser consecuencia de la existencia de fenotipos que
regulan la adaptación con más flexibilidad que otros; que poseen mejor
regulación de los procesos alostáticos minimizando los costes y aumentando los
beneficios (siguiendo el ejemplo del deportista profesional anterior,
ofreciéndole menor gasto fisiológico que sus adversarios, para similar nivel
competitivo o de entrenamiento) (Roelofs, 2010).
Esto se ve reflejado en los estudios de Alvero et al (2009), donde sobre una
muestra de jugadores de squash, se comparó la participación de la glucólisis
anaeróbica –controlando los niveles de lactato en sangre-, HR media y
percepción subjetiva de esfuerzo (RPE), entre W y L de encuentro de
competición. Los resultados arrojaron valores estadísticamente significativos
en los perdedores, que presentaban mayores niveles de lactato y RPE que los
ganadores. Lo que establecería relaciones directas entre manifestaciones
fisiológicas y L.
El
cortisol y la competición deportiva
Son muchos los estudios que se han desarrollado en materia de análisis de la
respuesta del C en la competición deportiva, por su relación con la
activación de los mecanismos de estrés (Tabla 1)
Tabla
1. Estudios sobre el impacto de la competición deportiva sobre los niveles
de C. (Adaptado de Salvador et al, 2005; 2009)
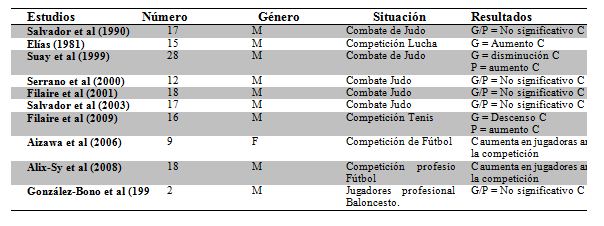
Los estudios son numerosos, pero inconclusos en cuanto a resultados, expresando
cambios enormes de unos deportes a otros, o de unos competidores a otros; lo
cual demuestra la idea de Robert Sapolsky sobre la respuesta neuroendocrina al
estrés: “Mayoritariamente ocurre, pero no siempre” (Sapolsky, 2004).
Entre los estudios que más nos interesaron para el presente trabajo, se
encuentran:
-
Filaire
et al (2009), sobre una muestra de 16 tenistas estudiando la respuesta
psicofisiológica al primer partido de tenis de una competición. Los
ganadores presentaron diferencias significativas con respecto a los
perdedores del primer partido; teniendo estos últimos más altos niveles de
C previos a la competición, durante la competición y basales al día
siguiente del partido. Los resultados de percepción subjetiva, también
presentaban diferencias significativas entre estos grupos, siendo nuevamente
los segundos quienes presentaban niveles más bajo de SC y mayores de CA,
así como en SA.
-
Mehta
et al (2008) apuntaron que la respuesta neuroendocrina al estatus social
competitivo, tenía relación con los niveles de T basales. Según Mehta, la
T es un buen precursor de los cambios en los niveles de C que acontecerán
en la competición deportiva; si la T basal es alta, la derrota deportiva
aumentará los niveles de C, mientras que si la T basal es baja, la derrota
o la victoria no tendrán una consecuencia significativa sobre los niveles
de C.
Estos estudios no están en desacuerdo con la HC, ya que son aquellos animales
que le dan importancia al recurso, los que generan T antes de las luchas;
aquellos que no muestran interés por el recurso (que no dan importancia a la
competición por el mismo) en el fondo ya han decidido, bien por indefensión
aprendida, bien por SEE, que los beneficios que puede obtener son claramente
insuficientes frente a los costes psicofisiológicos que le supone la lucha
intraespecífica.
La estrategia de Halcón (lucha-huída) presenta unos beneficios importantes, al
acceder al recurso de manera preferencial tanto alimentario como reproductivo,
sin embargo los costes son altos, ya que en las luchas continuas hay una alta
probabilidad de encontrarse con otro Halcón (H) más fuerte, que venza en ese
enfrentamiento, pudiendo llegar a infringir heridas graves e incluso la muerte
en su adversario.
La Estrategia de Paloma (D) por el contrario, no arriesga en cuestiones de
luchas por el recurso, pero en situación de escasez, las D sólo pueden optar
por mantener los niveles de reservas energéticas, pues velarán por su
supervivencia durante estas épocas.
La Estrategia Evolutivamente Estable, y la que aplican la mayoría de los
animales en libertad, es que cuando el recurso está disponible y es importante
para la procreación o la supervivencia del individuo, se lucha por él; por el
contrario, cuando el recurso en juego está en posesión de un tercero o su
relevancia evolutiva no es demasiada; los animales lanzarían señales
deshonestas con la intención de observar si el poseedor se las cree y abandona
el recurso voluntariamente o sin luchas importantes.
Cortisol
y estados de ánimo negativos
Muchos estudios han asociado el C con las diferencias individuales en los
estados de ánimo (Perna et al, 1998; Filaire et al, 2009), estados depresivos
(Burke et al, 2005; Stetler y Miller, 2005; Maximino et al, 2010; McNaugton y
Corr, 2004). En los estudios de Perna (1998), altos niveles de entrenamiento,
aumenta los niveles de C, correlacionando con las subescalas FATIGA y
DEPRESIÓN; deduciendo que el sobre-entrenamiento tiene una necesaria relación
con la activación del eje HPA.
Los resultados en competición, son importantes moduladores del aumento de C, y
de los estados de ánimo negativos y baja Autoconfianza (SC) (González-Bono et
al, 1999; Filaire et al, 1999; Alix-Sy et al, 2008).
Ekkekakis (2005), consideraba que el tipo de actividad física desarrollada,
ofrecerá una mejora de la SC y de los estados de ánimo; o en su defecto,
sensaciones desagradables físicas y psíquicas (Ekkekakis, 2007). Es lógico
preguntarse si existe relación entre el resultado final, y la percepción de
esfuerzo realizado. Si el afrontamiento ha sido activo y, pese al esfuerzo
físico, no se ha alcanzado la victoria; ¿provocaría el resultado un
desequilibrio momentáneo en el sistema neuroendocrino en aquellos deportistas
tras los desafíos deportivos? ¿Sólo ocurriría en los perdedores? ¿Son los
estados de ánimo negativos sistemas defensivos inhibidores de nuevas conductas
agonísticas?
Algunas
consideraciones sobre la transferencia de hormonas a la saliva
La saliva mantiene húmedo el aparato digestivo superior, favoreciendo la
ingesta de alimentos; sus enzimas ayudan la digestión, aporta una respuesta
inicial antibacteriana que inhibe la placa y también es un buen portador de
moléculas hormonales. Las capas lipofílicas de los capilares y las células
epiteliales transportan a la saliva moléculas, como por ejemplo esteroides, con
mayor rapidez que los péptidos.
Diferentes métodos analíticos han mejorado los procesos de medida hormonal
desde muestras de saliva. Los inmunológicos, son relativamente fáciles de
usar, los kits estandarizados se adquieren en el mercado a precios razonables y
los resultados son muy fiables. La cromatográfica es muy eficaz a la hora de
aportar datos cuantitativos. Las hormonas esteroides son importantes para el
control de las funciones del cuerpo humano: sistema neuronal, endocrino e
inmunológico.
Estas hormonas, analizadas como biomarcadores, han sido usadas en diagnósticos
clínicos de estrés (Nguyen y Wong, 2006), hiperplasia suprarrenal (Young et
al, 1988) o infertilidad; a nosotros nos interesan los controles en el ámbito
del deporte y de la medicina del deporte (Filaire et al, 2001, 2009; Salvador et
al, 2003). El uso de la saliva como medio diagnóstico, es un método muy
interesante para el futuro control de la actividad física y deportiva, porque
la recogida de muestras es rápida, sencilla y no reviste invasión alguna. Esto
y el hecho de que las hormonas esteroides no se unen a las proteínas en la
saliva, convierten este método en una excelente evaluación de las hormonas
esteroides libres presentes en la misma, antes, durante y tras la competición
deportiva (Plenis y Baczek, 2010).
El C es una de las hormonas más investigadas en saliva, como hemos visto, por
su relación con la gluconeogénesis en el estrés, y por tanto, de interés
para la investigación psicológica, como prueba de actividad adrenérgica
(Filaire et al, 2001, 2009; Perna et al, 1998; Serrano et al, 2000). El C es una
hormona esteroide procedente del córtex adrenal, como respuesta a la hormona
ACTH. Aumenta la presión arterial, los niveles de glúcidos, suprime el sistema
inmune; actúa sobre receptores específicos y tiene numerosas actuaciones en
algunos mecanismos psicológicos. La mayoría del C que encontramos en saliva
llega a través de mecanismos intracelulares.
La T es una hormona esteroide del grupo de los andrógenos y, al igual que el C,
ha sido ampliamente estudiada con éxito a través de la saliva, por su alta
correlación con la T circulante (Salvador, 2005, 2009). Los efectos de la T se
podrían clasificar como vigorizantes, con efectos anabolizantes como el aumento
de la masa muscular, de la densidad ósea, y cómo estimuladora del crecimiento
o desarrollo sexual en adolescentes.
En la actividad deportiva, el uso de técnicas de análisis hormonal ofrece un
respaldo científico importante, con unos protocolos de toma de muestras poco
invasivos y relativamente sencillos, lo que influye en una mayor participación
en estudios de campo en competición deportiva oficial. Esto ofrece un sistema
prometedor de control psicofisiológico de los deportistas de élite o promesas
deportivas en entrenamiento y competición (Edwards et al, 2006).
Conclusiones
Existen conductas más arriesgadas que otras para la consecución de un logro o
meta. El equilibrio es encontrar los beneficios y limitar los costes para
obtener los mejores resultados en la lucha, y la consecuente superación de los
competidores sociales. Este equilibrio satisface los niveles óptimos de
adaptación psicofisiológica y por tanto de los reajustes propios de la
supervivencia biológica. La diferencia entre la posición social relativa e
interés por la competición parece tener una relación con la serotonina, que
favorece la persistencia en la tarea y en la determinación de las conductas
arriesgadas (Hill y Buss, 2010).
La T se muestra como la gran protagonista en los procesos adaptativos en las
estrategias de supervivencia y adaptación. Son los individuos con mayores
niveles de esta hormona los que obtienen mejores resultados en la lucha; por el
contrario, la cortisolemia se relaciona con estados de ánimo negativos y el C
se encuentra presente en aquellos que pierden más partidos o están sometidos a
mayores niveles de exigencia psicofisiológica.
La toma de decisiones en los seres humanos puede cambiar a favor de los
resultados. Serán más arriesgadas cuando las habilidades cotidianas son
insuficientes para alcanzar la meta; lo que quiere decir, que inicialmente el
deportista analiza si las demandas fisiológicas y psicológicas se encuentran
dentro de las posibilidades de afrontamiento “seguro” (si es posible
alcanzar la meta sin conductas de riesgo), si no está dentro de sus
posibilidades de afrontamiento, el deportista decidirá si asume los riesgos que
supone un alto nivel de entrenamiento y competición deportiva, o asume una
conducta más conformista y conservadora. Parece ser que la T también juega su
papel, haciendo que los humanos con mayores niveles, asuman mayores conductas de
riesgo, que los que generan niveles más bajos, en situaciones de altas
exigencias competitivas.
En entrenamiento, sigue existiendo la idea de que entrenar el doble de tiempo,
implica proporciones similares de mejora psicofisiológica; sin embargo, niveles
de entrenamiento por encima de dichas posibilidades, puede desencadenar burnout
deportivo o graves problemas de salud; además de una inadecuación adaptativa.
Los deportistas expertos, son especialistas en la conservación de los recursos
energéticos y la optimización de su uso, mientras que los deportistas amateurs
son más impulsivos y nerviosos.
El estatus social podría manifestarse con relaciones lineales entre el ranking
deportivo y los niveles de T. Si esto fuera así, los jugadores que ostentan las
posiciones dominantes en la jerarquía están predispuestos a seguir manteniendo
esa posición jerárquica; los bajos niveles de C, los estados de ánimo
positivos, una positiva competencia percibida y la retroalimentación
psicofisiológica de la T, son poderosos procesos que envuelven y consolidan
esta idea.
Sin embargo, nuevos estudios (Mehta y Josephs , 2010, Liening y Josephs, 2010)
apuntan a una activación conjunta de ambos ejes, cuando de búsqueda de
conducta social se habla, lo cual abre una nueva línea de investigación que
ofrece una alternativa muy interesante a la visión inicial de la respuesta
psicofisiológica a la competición deportiva y social.
En estos estudios, publicados en Hormones & Behavior y otras revistas
de impacto, estos autores ofrecen la base científica neuroendocrina en la cual
basan su teoría de activación conjunta de ambos ejes, en la inhibición o
materia de búsqueda de estatus social. En los estudios desarrollados por Mehta
(2010), podemos observar como existe una relación significativa entre altos
niveles de T y bajos niveles de C, en aquellos deportistas que ganan más
encuentros, y que tienen mayor interés por nuevos encuentros agonísticos;
mientras que altos niveles de T y altos de C producen cambios muy marcados tras
la derrota y desinterés por nuevas conductas competitivas. Para Liening y
Josephs (2010), existen una serie de moderadores que ofrecen una clara
influencia sobre la T, como pueden ser la vasopresina en el caso del género
masculino, o el estradiol en el caso del género femenino. Para Josephs, la T
fomenta el deseo de estatus social, pero para que la conducta motivada de
aproximación en la búsqueda de estatus social se pueda desarrollar, bajos
niveles de C son necesarios. Esta Hipótesis se basa en la idea, apuntada por
Viau (2002), que indica la relación antagonista entre los ejes HPG e HPA,
cuando uno se activa el otro es suprimido; por tanto, un aumento del C en
respuesta a una situación de estrés competitivo o precompetitivo respondería
suprimiendo los efectos de la T en la búsqueda de estatus social o deportivo.
Bibliografía
-
Abbot,
D., Keverne, E., Bercovitch, F., Shievely, C., Mendoza, S., Saltzman, W, et
al. (2003) Are subordinates always stressed? A comparative analysis of rank
differences in cortisol levels among primates. Hormones and Behavior 43,
67-82.
-
Alix-Sy,
D., Le Scanff, C. y Filaire, E. (2008) Psychophysiological response in the
pre-competition period in elite soccer players. Journal
of Sport Science and Medicine 7, 446-454.
-
Alvero
Cruz, J.R.., Barrera, J., Mesa, A. y Cabello D. (2009) Correlation of
physiological responses in squash players during competition. 64-69 In:
Science and Racket Sports IV. Routledge. New York.
-
Archer,
J. (2004) Sex differences in aggression in real-world setting: A
meta-analitic review. Review of General Psychology, 8, 291-322.
-
Archer,
J. (2006) Testosterone and human aggression: an evaluation of the challenge
hypothesis. Neuroscience and
Behavioral Reviews 30 (3), 319-345.
-
Archer,
J.; Crahan-kevan, N. y Davies, M. (2005) Testosterone and aggression: a
re-analysis of book: Stazyk and Kinsey´s (2001). Study
of Aggressive and Violente Behavior, 10, 241-261.
-
Balaguer,
I., Fuentes, I., Meliá, J.L. y García-Merita, M.L. (1993) El perfil de los
estados de ánimo (POMS): Baremo para estudiantes valencianos y su
aplicación en el contexto deportivo. Revista
de Psicología del Deporte, 12, 49-68.
-
Bandura,
A. (1982) Self-efficacy mechanisms in human agency. American
Psychology 37, 122-147.
-
Bandura,
A (1997) Self-efficacy: the exercise of control. New York: W.H.
Freesman.
-
Bateup,
H.S., Booth, A., Shirtcliff, E.A. Granger, D.A. (2002) Testosterone,
cortisol and
-
Booth,
A. Shelley, G. Marzur, A., Tharp, G., Kittok, R. (1989) Testosterone and
winning and losing in human competition. Hormones
and Behavior, 23, 4, 556-571.
-
Bower,
J., Low, C. Moscowitz, J., Sepah, S., y Epel, E. (2008) Benefit finding and
physical health: positive psychological changes and enhanced allostasis. Social
and Personality Psychology Compass: 223-244.
-
Buchanan,
K., Evans, M., Goldsmith, A., Bryant, D., Rowe, L. (2001) Testosterone
influencies basal metabolic rate in male house sparrows: a new cost of
dominance?. Proc. Royal Society of
London 268, 1337-1344.
-
Buchanan,
K., Evans, M.R., Roberts, M. (2010) Does testosterone determine dominance in
the house sparrow? An experimental test. Journal
of Avian Biology, 41, 4, 445-451.
-
Buchanan,
T.W., Driscoll, D., Mowrer, S., Sollers, J., Thayer, J., Krischbaum, C.,
Tranel, D. (2010) Medial prefrontal cortex damage effects physiological and
psychological stress responses differently in men and women. Psychoneuroendocrinology
35, 56-66.
-
Carney,
D. y Manson, M. (2010) Decision making and testosterone: when the ends
justify the means. Journal or
Experimental Social Psychology 46, 668-671.
-
Carré,
J.M. y McCormick, C.M. (2008) Aggressive behavior and change in salivary
testosterone concentrations predict willingness to engage in a competition
task. Hormones and Behavior 54, 3,
403-409.
-
Carré,
J.M. y Putnam, S. (2010) Watching previous victory produce an increase in
testosterone among elite hockey players. Psychoneuroendocrinology,
35, 3, 475-479.
-
Carré,
J.M., Gilchrist, J.D., Morrissey, M. Y McCormick, C. (2010) Motivational and
situational factors and the relationship between testosterone dynamics and
human aggression during competition. Biological
Psychology, 84, 2, 346-353.
-
Cashdan,
E. (2003) Hormones and competition aggression in women. Aggressive
Behavior, 29, 2, 107-115.
-
Cladue,
B.A., Boechler, M. y McCoul, K.D. (1989) Hormonal response to competition in
human. Aggressive Behavior, 15,
409-422.
-
Clutton-Brock,
T. y Parker, G. (1995) Punishment in animal societies. Nature
373, 209-216.
-
Craig,
I.W., Halton, K.E. (2009) Genetics of human aggressive behavior. Human
Genetic 126, 101-113.
-
Creel,
S. (2001) Social dominance and stress hormones. Trends
in Ecology and Evolution, 16, 439-497.
-
Crewther,
B.T. y Cook, C. (2010) Measuring the salivary testosterone and cortisol
concentrations of weighlifters using the enzyme-immunoassay kit. International
Journal of Sports medicine, 7, 486-489.
-
Crewther,
B.T., Lowe, T.E., Ingram, J. y Weatherby, R.P. (2010) Validating the
salivary testosterone and cortisol concentration measures in response to
short high-intensity exercise. Journal
of Sport Medicine Physiology and Fitness, 50, 1, 85-92.
-
Dorn,
L.D., Kolko, D.J., Susman, E.J., Huang, B., Stein, H., Music, E. et al.
(2009) Salivary gonadal and adrenal hormone differences in boys and girls
with and without disruptive behavior disorders: contextual variants. Biological
Psychology, 81, 1, 31-39.
-
Durban,
R. (2003) The social brain: mind, language, and society in evolutionary
perspective. Annual Review of
Anthropology, 32, 163-181.
-
Ebstein,
R., Israel, S., Chew, S., Zhong, S., Knafo, A. (2010) Genetics of human
social behavior. Neuron Reviews 65,
831-844.
-
Edwards,
D.A. y Kurlander, L.S. (2010) Women´s intercollegiate volleyball and
tennis: effects of warm-up, competition and practice on saliva levels of
cortisol and testosterone. Hormones
and Behavior 58, 4, 606-613.
-
Edwards,
D.A., Wetzel, K., Wyner, D.R. (2006) Intercollegiate soccer: saliva cortisol
and testosterone are elevated during competition, and testosterone is
related to status and social connectedness with team mates. Physiology
and Behavior 87:135-143
-
Ekkekakis,
P. (2003) Pleasure and displeasure from the body: perspectives from
exercise. Cognition and Emotion 17
(2), 213-239.
-
Ekkekakis,
P., Eric, H., Petruezzello, J. (2005) Variation and homogeneity in affective
responses to physical activity of varying intensities: an alternative
perspective on dose-response based on evolutionary considerations. Journal
of Sports Sciences 23, 5, 477-500
-
Ekkekakis,
P. (2008) Affect circumplex redux: the discussion on its utility as a
measurement framework in exercise psychology continues. International
Review of Sports and Exercise Psychology 1, 2,139-159.
-
Ekkekakis,
P. (2009). The dual-mode theory of affective responses to exercise in
metatheoretical context: initial impetus, basic postulates and philosophical
framework. International Review of
Sport and Exercise Psychology 2, 1, 73-94.
-
Eriksen,
H. Murison, R. Pensgaard, A. y Ursin, Holger (2005) Cognitive activation
theory of stress (CATS): From Fish brains to the Olympics. Pyschoneuroendocrynology
30, 933-938
-
Eriksen,
H. Ursin, H. (2002) Social inequalities in health: biological, cognitive and
learning theory perspective. Norsk
Epidemiology 12, 33-38
-
Epstein,
L., Cluss, P. (1986) Behavioral genetics of childhood obesity. Behavior
Therapy, 17, 4, 324-334.
-
Filaire,
E., Alix, D., Ferrand, C., Verger, M. (2009) Psychophysiological stress in
tennis players during the first single of a tournament. Psychoneuroendocrinology,
34, 150-157.
-
Filaire,
E., Maso, F., Sagnol, M., Lac, G. (2001) Anxiety, hormonal responses, and
coping during a judo competition. Aggressive
Behavior 27, 55-63.
-
Filaire,
E., Sagnol, M., Ferrand, C., Maso, F. y Lac. G. (2001) Psychophysiological
stress in judo atheletes during competitions. Journal
of Sports Medicine and Physical Fitness 41, 2, 263-268.
-
Girard,
O., Millet, G. (2008) Neuromuscular fatigue in racquet sports. Neurologic
Clinics 26, 181-194.
-
Girard,
O., Sciberras, P., Habrard, M., Hot, P., Chevalier, R., Millet, G.P. (2005)
Specific incremental test in elite squash players. British
Journal of Sports Medicine 39, 921-926.
-
González-Bono,
E., Salvador, A., Serrano, M.A., Ricarte, J. (1999) Testosterone, cortisol
and mood in a sport team competition. Hormones
and Behavior 35, 55-62.
-
Gozansky,
W.S.; Lynn, J.S.; Laudenslager, M.L.; Kohrt, W.M. (2005) Salivary cortisol
determined by enzyme immunoassay is preferable to serum total cortisol for
assessment of dynamic hypothalamic-pituitary-adrenal axis activity. Clinical
Endocrinology 63, 3, 336-341.
-
Gröschi,
M. (2008) Current status of salivary hormone analysis. Clinical Chemistry
54, 11, 1759-1769.
-
Haller,
J., Van de Schraaf, J., Kruk, M. (2001) Deviant forms of aggression in
glucocorticoid hyporeactive rats: a model for pathological aggression? Journal
of Neuroendocrinology 13, 102-107.
-
Hanershi,
K., Fry, A., Moore, C.A., Schilling, B.K., Li, Y., Fry, M.D. (2007) Cortisol
and stress responses during a game and practice in female collegiate soccer
players. Journal of Strength and
Conditioning Research 21, 2, 583-588.
-
Haren,
M.T., Siddiqui, A.M., Ambrecht, H.J., Kevorkian, R.T., Kim, M.J., Haas,
M.J., et al (2010) Testosterone modulates gene expression pathways
regulating nutrient accumulation, glucose metabolism and protein turnover in
mouse skeletal muscle. International
Journal of Andrology, Article. In Press.
-
Harvey,
A.H.; Siperstein, A.A. y Berber, E. (2010) Clinical detection and treatment
of adrenal disease. Molecular
Pathology of Endocrine Diseases, 3, 5, 197-203.
-
Hermans,
E., Putman, P., Baas, J., Koppeschaar, H., van Honk, J. (2006) Single
administration of testosterone reduces fear-potentiated startle in humans. Biological
Psychiatry, 59, 872-874.
-
Hill,
S.E., Buss, D.M. (2010) Risk and relative social Rank: positional concerns
and risk shifts in probabilistic decision-making. Evolution
and Human Behavior 31, 219-226.
-
Jiménez,
M. (2010) El badminton: una revisión actual. EFdeportes.
Revista Digial nº 149.
-
Jiménez,
M. y Videra, A. (2010) Estudio de la ansiedad precompetitiva, los estados de
ánimo y su relación con el Burnout en jugadores de bádminton de liga
nacional de clubes de división de honor. EFDeportes.com, Revista
Digital. Buenos Aires, nº 149. http://www.efdeportes.com/efd149/estudio-de-la-ansiedad-y-burnout-en-badminton.htm
-
Johnson,
R.T., Burk, J.A., Kirkpatrick, L.A. (2007) Dominance and prestige as
differential predictors of aggression and testosterone levels in men. Evolution
and Human Behavior, 28, 5, 345-351.
-
Kiulighan,
K.T., Granger, D.A. y Booth, A. (2004) Gender differences in testosterone
and cortisol response to competition. Psychoneuroendocrinology
30, 1, 58-71.
-
Korte,
S., Koolhaas, J.M., Wingfield, J, McEwen, B. (2005). The darwinian concept
of stress: benefits of allostais and cost of allostasis load and the trade
off in health and disease. Neurocience
and Behavioral Reviews 29, 3-38.
-
Korte,
S., Oliver, B., Koolhaas, J. (2006) A new welfare concept base don
allostasis. Physiology and Behavior, 92, 422-428.
-
Levine
S. y Ursin, H. (1991) What´s is stress? Stress: neurobiology and
neuroendocrinology. Marcel Dekker. Nueva York. 3-21
-
Lupien,
S., McEwen, B., Gunnar, M., Heim, C. (2009) Effects of stress thoughoutnthe
lifespan on the brain, behaviour and cognition. Nature,
10, 434-445.
-
Manning,
J, Bundred, P., Newton, D., Flanagan, B. (2003) The second or fourth digit
ratio and variation in the androgen receptor gene. Evolution
and Human Behavior, 24, 399-405.
-
Martens,
R.; Vealey, R, Burton, D. (1990) Competitive Anxiety in Sport. Human
Kinetics. Illinois.
-
Martinez,
A., Seco, J., Tur, M., Josep, A., Abecia, L., Orellana, E., Biescas, A.
(2010) Testosterone and cortisol changes in professional basketball players
though a season competition. Journal
of Strength and Conditioning Research 24, 4, 1102-1108.
-
Marzur,
A. (1985) A biosocial model of status in face to face primate groups. Social
Forces, 64, 377-402.
-
Marzur,
A., Booth, A., Daqbbs, J.M. (1992) testosterone and chess competition. Social
Psychology Quarterly, 55, 1, 70-77.
-
Marzur,
A., Lamb, T. (1980) Testosterone, status and mood in human males. Hormones
and Behavior, 14, 236-246.
-
Marzur,
A. y Booth, A. (1998) Testosterone and dominance in men. Behavior
Brain Science, 21, 353-397.
-
Mason,
J.W. (1968) A review of psychoendocrine research on the pituitary-adrenal
cortical system. Psychosomatic Medicine 30, 576-607.
-
Maximino,
C., Marques de Brito, T., Gouveia, A. (2010) Construct validity of
behavioral model of anxiety: where experimental psychopathology metes
ecology and evolution. Psychology and
Neuroscience, 3, 1, 1-25.
-
Mehta,
P., Jones, A., Josephs, R. (2008) The social endocrinology of dominance:
basal testosterone predicts cortisol changes and behavior following victory
and defeat. Journal of Personality
and Social Psychology 94, 6, 1078-1093.
-
Mehta,
P., Wuehrmann, E.V., Joseph, R.A. (2009) When are low testosterone levels
advantagenus? The modelating role of individual versus intergroup
competition. Hormones and Behaivor 56, 158-162.
-
McAndrew,
F.T. (2009) The interacting roles of testosterone and challenge to status in
human male aggression. Aggression and
Violent Behavior, 14, 5, 330-335.
-
McBurnett,
K., Lahey, B.B., Rathouz, P.J. y Loeber, R. (2000) Low salivary cortisol and
persistent aggression in boys referred for disruptive behavior. Archives
of General Psychiatry, 57, 38-43.
-
McBurnett,
K., Raine, A., Stouthamer-Loeber, M., Loeber, R., Kumar, A.M., Kumar, M.
(2005) Mood and hormones responses to psychological challenge in adolescent
male with conduct problems. Biological
Psychiatry, 57, 1109-1116.
-
McEwen,
B. (1998) Stress, adaptation, and disease. Allostasis and allostasis load. Annual
New York Academic of Science, 840, 33-44.
-
McEwen,
B. y Wingfield, J. (2003). The concept of allostasis in biology and
biomedicine. Hormones and Behavior Nº 43, 2-15.
-
McEwen,
B. y Wingfield, J. (2010). What´s is in name? Integrating homeostasis,
allostasis and stress. Hormones and
Behavior, 57, 105-111.
-
Mc
Grath (1970) A conceptual formulation for research on stress. Social and
psychological factor in stress. Rinehart and Winston. Nueva York
-
McKinney,
W.T. (1984). Animal models of depression: an overview. Psychiatric
Development, 2, 77-96
-
McNair,
D.; Lorr, M.; Droppleman, L. (1971) Profile of mood states manual.
Educational and industrial testing services. California.
-
McNaughton,
N. y Corr, P.J. (2004) A two-dimensional neuropsychology of defense:
fear/anxiety and defensive distance. Neuroscience
and Biobehavioral Revivews, 28, 285-305.
-
Medina,
G. y García, F. (2002) Burnout, locus de control y deportistas de alto
rendimiento. Cuadernos de Psicología
del Deporte, II, 2, 29-42.
-
Moreira,
A., Assati, F., Oliveira, Y.B., da Silva, D.A., de Araujo, V.C. (2009)
Salivary cortisol in top-level proffesional soccer players. European
Journal of Applied Physiology, 106,1, 25-30.
-
Moya-Albiol,
L., Salvador, A., Costa, R., Ferrand, G., Lac, G. (2001) Anxiety, hormonal
responses and coping during a judo competition. Aggressive
Behavior, 27,1, 55-63.
-
Muller,
N. Wranghan, R. (2004) Dominance, aggression and testosterone in wild
chimpanzees: a test of the challenge hypothesis.
Annual Behavior 67, 113-123.
-
Nowak,
A., Straburyriska-lupa, A., Kusy, K., Zielinski, d., Ritwenger, J.
Karolkiewicz, E. (2010) Bone mineral density and bones turnover in male
masters athletes aged 40-64. The
Aging Male, 13, 2, 133-141.
-
Ntoumanis,
N. y Jones, G. (1998) interpretation of competitive trait anxiety symptoms
as a function of locus of control beliefs. International
Journal of Sport Psychology. 29, 99-114.
-
Oliveira,
T., Gouveira, M.J., Oliveira, R.F. (2009) Testosterone responsiveness to
winning and losing experiencies in female soccer players. Psychoneuroendocrinology
34, 7, 1056-1064.
-
Ontony,
A. y Turner, J. (1990) What´s Basic about basic emotion?. Psychological
Review 3, 315-331.
-
Oxford,
J., Ponzi, D. Geary, D.C. (2009) Hormonal response differ when playing
violent videogames against an ingoup and outgroup. Evolution
and Human Behavior 31, 3, 201-209.
-
Pensgaard,
A.M., Roberts, G.C. (2003) Achievement goal orientations and use of coping
strategies among winter Olympians. Psychology
and Sport Exercises 4, 1001-1116.
-
Perez-Rodriguez,
L. Blas, J. Viñuela, J.; Marchant, T.A.; Bortolotti, G.R. (2006) Condition
and androgen levels: are condition-dependent and testosterone-mediated
traits two sides of the same coin? Animal
Behavior 72, 97-103.
-
Perna,
F., Antoni, M., Kumar, M., Cruess, D. y Schneiderman, N. (1998)
Cognitive-behavioral intervention effects on mood and cortisol during
exercise trainings. Annual Behavioral
Medicine 20, 2, 92-98.
-
Plenis,
A. y Baczek, T. (2010). Quantification of salivary steroid hormones
considered as bio-markers in clinical research studies and sport medicine. Current
Pharmaceutical Analysis, 6, 3, 182-197.
-
Quinn,
J.F. y Dunham, A.E. (1983) On hypothesis testing in ecology and evolution. American
Naturalist 122, 602-617.
-
Rohwer,
S.A. y Ewald, P.W. (1981) The cost of dominance and advantage of
subordination in a badge signaling system. Evolution
35, 441-454.
-
Rolls,
E.T. (2005) Emotion explained. Oxford University Press. Oxford.
-
Roy,
M., Krischbaum, C., Dteptoe, A. (2003) Intraindividual variation in recent
stress exposure as a moderator of cortisol and testosterone levels. Annual
of Behavioral Medicine 26, 3, 194-200.
-
Ruiz,
J., Mesa, J., Mingorance, I., Rodríguez-Cuartero, A., Castillo, M. (2004)
Deportes con alto grado de estrés físico afectan negativamente al perfil
lipídico plasmático. Revista
Española de Cardiología, 57, 6, 499-506.
-
Sachser,
N. y Kaiser, S (2010) The social modulation of behavioural development.
Animal Behavior: Evolution and Mechanisms. Springer. Berlin
-
Salvador,
A., Suay, F., Martínez-Sanchís, S., González-Bono, E. (1995) Deporte y
salud : efectos de la actividad física sobre el bienestar psicológico
y mecanismos hormonales subyacentes. Revista
de Psicología General y Aplicada 48, 1, 125-137.
-
Salvador,
A., Suay, F., González-Bono, E., Serrano, M.A. (2003) Anticipatory
cortisol, testosterone & psychological responses to judo competition in
young men. Psychoneuroendocrinology
28, 364-375
-
Salvador,
A. (2005) Coping with competitive situations in humans. Neuroscience
and Biobehavioral Reviews 29, 195-205.
-
Salvador,
A y Costa, R. (2009) Coping with competition: Neuroendocrine responses and
cognitive variables. Neuroscience and Biobehavioral Reviews 33, 160-170.
-
Sapolsky,
R. (2004) Social status and health in humans and other animals. Annual
Review of Anthopology 33, 393-418.
-
Sapolsky,
R. (1992) Neuroendocrinology of the stress response. Behavioral
Endocrinology, 287-324.
-
Sapolsky,
R. (1991) Testicular function, social rank, and personality among wild
baboons. Psychoneuroendocrinology 16,
281-293.
-
Senar,
J.C., Polo, V., Uribe, F., Camerino, M. (2000) Status signalling, metabolic
rate and body mass in the siskin: the cost of being a subordinate. Animal
Behavior, 59, 103-110.
-
Serrano,
M., Salvador, A., González-Bono, Sanchís, C., Suay, F. (2000) Hormonal
response to competition. Psicothema,
12, 3, 440-444.
-
Seyle,
H. (1936) A syndrome producted by diverse nocuous agents. Nature
138, 32.
-
Seyle,
H. (1950) Stress and general adaptation syndrome. British
Journal of Medicine 1, 4667, 1383-1392.
-
Seyle,
H. (1956) The Stress of Life. McGrawnHill. New York
-
Seyle,
H. (1974) Stress without Distress. Lippicott. Philadelphia
-
Shoal,
G.D., Giancola, P.R. y Kirillova, G.P. (2003) Salivary cortisol,
personality, and aggressive behavior in adolescent boys: a 5-year
longitudinal study. Journal of the
American Academy of Child and Adolescent psychiatry, 42, 9, 1101-1107.
-
Sinervo,
B., Clobert, J., Miles, D.B., McAdam, A., Lancaster, L.T. (2008) The role of
pleiotropy vs signaler-receiver gene epistasis in life history trede-offs:
dissecting the genomic architecture of organismal design in social systems. Heredity
101, 197-211.
-
Smekal,
G., Von Duvillard, S., Rihacek, C., Pokan, R., Hofmann, R., Tschan, H.
Bachl, N. (2000) A physiological profile of tennis match play. Journal
of Medicine and Science in Sports and Exercise, 999-1005.
-
Smiley-Oyen,
A., Lowry, K., Francois, S., Kohut, M., Ekkekakis, P. (2008) Exercise,
Fitness, and neurocognitive function in older adults: the “selective
improvement” and “cardiovascular fitness” hypotheses. Annual
Behavioral Medicine 36, 280-291.
-
Smith,
E.A. (2010) Comunication and collective action: lenguaje and the evolution
of human cooperation. Evolution and
Human Behavior, 31, 4, 231-245.
-
Smith,
M. (1974) The Theory of Games and the evolution of animal conflicts. Journal
of TheoricaL Biology 47, 209-221.
-
Smith,
M. (1984) Game theory and the evolution of behavior. Behavioral and Brain
Sciences, 7, 95-101.
-
Spielberger,
C.D. (1972) Conceptual and methodological issues in anxiety research. Anxiety:
Current Trends in Theory and Research. Academic press. New York.
-
Stetler,
C. y Miller, G.E. (2005) Blunted cortisol response to awakening in mild to
moderate depression: regulatory influences of sleep patterns and social
xontacts. Journal of Abnornal
Psychology, 114, 687-705.
-
Suay,
F., Salvador, A., González-Bono, E., Sanchis, C., Martínez, M., Martínez,
S., Simon, V.M. y Montoro, J.B. (1999) Effects of competition and its
outcome on serun testosterone, cortisol and prolactin. Psychoneuroendocrinology,
24, 5, 551-556.
-
Susman,
E.J., Dockray, s., Schiefelbein, V.L., Herwehe, S., Heaton, J.A. y Dorn,
L.D. (2007) Morningness/eveningness, morning-to-afternoom cortisol ratio,
and antisocial behavior problems during puberty. Development
Psychology, 43, 4, 811-822.
-
Szczepanska-sadowska,
E. (2008) Role of neuropeptides in central control of cardiovascular
response to stress. Journal of
Physiology and Pharmacology 59, 8, 61-89.
-
Tausk,
M. (1951) Hat die nebenniere tatsächlich eine verteidingungsfunktion?
Organon. Holland
-
Thompson,
R., Mata, J., Jaeggi, S., Buschkuehl, M., Jonides, J. (2010) Maladaptative
coping, adaptative coping, and depressive symptoms: variations across age
and depressive state. Behaviour
Research and Therapy 48, 6, 459-466
-
Tomassini,
M., Pestelacci, E., Luthi, L. (2010) Mutual trust and cooperation in the
evolutionary haws-doves game. Biosystems
99, 1, 50-59.
-
Trenayne,
P. y Barry, R.J. (2001) Elite pistols shooters: physiological patterning of
best vs worst shots. International
Journal of Psychophysiology, 41, 1, 19-29.
-
Urhausen,
A., Gabriel, H., Kendarmanny, W. (1995) Blood hormones and markers of
training stress and overtraining. Journal
of Sports Medicine 20, 4, 251-276.
-
Ursin,
H. y Eriksen, H.R. (2004) The cognitive activation theory of stress. Psychoneuroendocrinology
29, 567_592.
-
Van
Anders, S.M., Watson, N.V. (2006) Social neuroendocrinology. Effects of
social contexts and behaviors on sex steroids in humans. Human
Nature 17, 212-237.
-
Van
Anders, S.M., Watson, N.V. (2007) Effects of ability and change determined
competition outcome on testosterone. Physiological
Behavior 90, 634-642.
-
Van
Bokhoven, I., van Goozen, S.H., van Engeland, H., Schaal, B., Arseneault,
L., Séguin, J.R., et al. (2005) Salivary cortisol and aggression in a
population-based longitudinal study of adolescent males. Journal
of Neural Transmission, 112, 8, 1083-1096.
-
Van
der Meij, L., Buunk, A., Almela, M. y Salvador, A. (2010) Testosterone
responses to competition: the opponent´s psychology state makes it
challenging. Biological Psychology
84, 2, 330-335.
-
Veissier,
I., Boissy, A. (2007) Stress and welfare: two complementary concepts that
are intrinsically related to the animal´s point of view. Physiology
and Behavior 92, 479-433.
-
Vinnig,
R.F., McGinley, R.A., Symons, R.G. (1983) Hormones in saliva: mode of entry
and consequent implications for clinical interpretation. Clinical
Chemistry 29, 1752-1756.
-
Vreeburg,
S.A.; Zitman, F.G.; Van Pelt, J. DeRijk, R.H.; Verhagen, J.; Van Dyck, R.;
Hoogendijk, J.G.; Smith, J. Brenda, W.J. (2010) Salivary cortisol levels in
persons with and without different anxiety disorders. Psychosomatic
Medicine 72, 340-347.
-
Wagner,
J.D., Flinn, M.V. y England, B.G. (2002) Hormonal response to competition
among male coalitions. Evolution and
Human Behavior, 23, 6, 437-442.
-
Wingfield,
J.C. (2005) The concept of allostasis: coping with a capricious environment.
Journal of Mammalogy, 86, 2, 248-254.
-
Wingfield,
J.C., Hegner, R., Dufty, A., y Ball, G. (1990) The challenge hypothesis:
Theoretical implications for patterns of testosterone secretion, mating
systems, and breeding strategies. American
Nature 136, 829-846.
-
Wingfield,
J.C., Lynn, S., Sona, K. (2001) Avoiding the cost of testosterone:
ecological bases of hormone-bahavior interactions. Brain
Behavior Evolution 57, 239-251.
-
Wüst,
S., Federenko, I.S., Van Rossum, E.F., Koper, J., Hellhammer, D.H. (2005)
Habituation of cortisol responses to repeated psychosocial stress-futher
characterization and impact of genetic factors. Psychoneuroendocrinology
30, 199-211.
-
Young,
M.C., Walker, R.F., Riad Fahmy, D., Hughes, I.A. (1988) Androstenedione
rhythms in saliva in congenital adrenal hyperplasia. Archives
of Childhood, 63, 624-628.
Otros artículos sobre Entrenamiento
deportivo

