
|
|||
|---|---|---|---|
|
|
Flexibilidade: aspectos fisiológicos e fatores limitantes La flexibilidad: aspectos fisiológicos y factores limitantes |
|
|
|
Laboratório de Pesquisa do Exercício, LAPEX Universidade Federal do Rio Grande do Sul, Porto Alegre (Brasil) |
Marcos Franken |
|
|
|
Resumo A flexibilidade pode ser definida como a máxima amplitude fisiológica passiva em um determinado movimento articular necessário que irá depender da elasticidade muscular e da mobilidade articular, sem que ocorram lesões anátomo – patológicas. Pode ser influenciada pelo sexo, idade, o treinamento, raça, histórico de lesão, reflexos espinhais, a temperatura, a hora do dia e a composição do músculo. Dois mecanorreceptores importantes são os fusos musculares e os órgãos tendinosos de Golgi, e os receptores articulares sentem forças mecânicas nas articulações como a pressão de alongamento e distensão. O sistema nervoso funciona por meio de um conjunto de interações denominados reflexos, como o reflexo de alongamento, a inibição recíproca e o reflexo miotático inverso. Além do mais, a co-contração tem sido estabelecida durante vários movimentos a nível articular. O tecido conjuntivo é influenciado pelo envelhecimento, imobilização, distúrbios metabólicos e deficiências nutricionais, e por ser um dos componentes mais influentes na limitação da amplitude do movimento, devem ser bem alongados. Unitermos: Flexibilidade. Aspectos fisiológicos. Exercício.
|
|||
|
|
EFDeportes.com, Revista Digital. Buenos Aires, Año 15, Nº 148, Septiembre de 2010. http://www.efdeportes.com/ |
|
|
1 / 1
Introdução
A flexibilidade é uma das variáveis essenciais da aptidão física relacionada a saúde, sendo definida como a máxima amplitude fisiológica passiva em um determinado movimento articular necessário que irá depender da elasticidade muscular e da mobilidade articular, sem que ocorram lesões anátomo – patológicas (ALTER, 1999). Juntamente com a força, a resistência, a potência, o equilíbrio e a coordenação, a amplitude de movimento contribui de modo significativo para a qualidade global das funções físicas de um indivíduo. A amplitude passiva de um movimento articular depende primariamente da estrutura e da função do osso, do músculo e do tecido conectivo e de outros fatores tais como o desconforto e a habilidade para gerar força e potência muscular suficiente, para isso é importante que se conheça os aspectos fisiológicos que irão interferir na amplitude do movimento. Este artigo de revisão bibliográfica tem como objetivo descrever os aspectos fisiológicos da flexibilidade e os fatores limitantes, que são importantes para o entendimento deste componente da aptidão física.
Aspectos fisiológicos da flexibilidade
Para executar um trabalho de alongamento dos músculos efetivo, devemos conhecer as propriedades neurofisiológicas dos músculos que podem afetar um aumento da flexibilidade. Para isso, três receptores principais tem implicações para o alongamento e a manutenção da amplitude de movimento favorável, como o fuso muscular, o órgão tendinoso de Golgi (OTG) e os mecanorreceptores articulares e descritos quatro fenômenos neurofisiológicos importantes, como o reflexo miotático, a coativação/co-contração, inibição autógena e a inibição recíproca (BANDY & SANDERS, 2003).
Fuso muscular
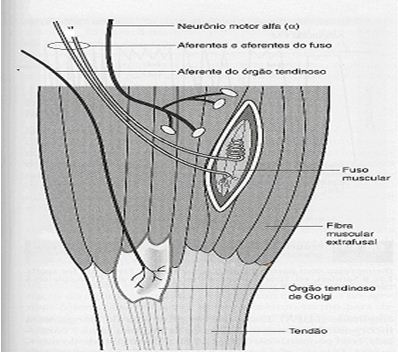
O fuso muscular é um receptor que consiste de fibras musculares especiais, como as fibras intrafusais que são distintas das fibras musculares esqueléticas normais (fibras extrafusais) (Figura 1), terminações sensoriais e motoras localizadas nos músculos, tendões e vestíbulo da orelha cujo reflexo está conectado com a locomoção ou a postura (Gordon & Ghez, 1991; Alter, 1999). São numerosos principalmente nos músculos pequenos e delicados da mão e do olho e é o principal órgão sensitivo do músculo.
Alterações no comprimento do músculo e à velocidade com que o comprimento se altera irão influenciar as terminações sensoriais. No alongamento do músculo, as fibras intrafusais estendem e fazem contato com as fibras extrafusais (BANDY & SANDERS, 2003). O processo de excitação de um músculo acontece quando uma mudança na permeabilidade da extremidade do neurônio sensorial ocorre, resultando em uma transferência de carga por meio da membrana terminal do nervo. Esse fluxo de corrente produz uma despolarização, denominada potencial gerador. Quando a despolarização alcança o limiar, resulta em um potencial de ação conduzido. O fuso muscular mantém um feedback contínuo de informações de cada músculo ao sistema nervoso central, apresentando a situação do músculo em cada momento (Guyton, 1996).
Os fusos musculares são compostos de três tipos de fibras intrafusais (bolsa nuclear dinâmica, bolsa nuclear estática e cadeia nuclear), que se unem para funcionar como fusos primários e secundários. Os fusos primários são influenciados pela taxa de mudança no comprimento, criando uma resposta dinâmica. Os fusos secundários são influenciados por uma mudança no comprimento estático absoluto, ou seja, quando o estiramento muscular ativa o reflexo do fuso, as fibras extrafusais contraem-se encurtando o músculo (FRONTERA et al., 2001).
Órgão Tendinoso de Golgi
O OTG é um mecanoreceptor sensível à contração dos músculos esquelético, a pequenas alterações na tensão do tendão e responde a tensão adicional tanto por estiramento passivo de um músculo como por contração muscular ativa (BANDY & SANDERS, 2003; ALTER, 1999). São localizados nas aponeuroses, ou junções músculo-tendinosas, e não dentro dos tendões (Figura 1).
A principal tarefa do OTG é impedir a atividade excessiva das fibras nervosas que inervam o músculo extrafusal. Se os músculos forem alongados por 15 a 30 segundos em toda a sua extensão, acarreta em tensão no tendão. O OTG responde a essa tensão por meio das fibras nervosas tipo Ib, sendo que essas fibras nervosas têm a capacidade de sobrepor os impulsos provenientes do fuso muscular, acarretando em um relaxamento reflexo. Sendo assim, os músculos relaxam e conseguem alongar-se (Gordon & Ghez, 1991). Sua sensibilidade é tão grande que eles podem responder à contração de uma única fibra muscular (Wilmore & Costill, 2001).
Percebeu-se que o OTG exibe um limiar mais baixo à contração muscular que ao alongamento passivo do músculo devido a força da contração muscular ser transferida de forma mais eficaz ao OTG do que a força do alongamento muscular (Monteiro, 2006).
Figura 1. Fuso Muscular e Órgão Tendinoso de Golgi (adaptado de BANDY & SANDERS, 2003).
Mecanorreceptores articulares
Todas as articulações sinoviais do corpo humano contem quatro receptores de extremidades nervosas. Esses receptores articulares sentem forças mecânicas nas articulações, tais como pressão de alongamento e distensão. Esses receptores podem ser classificados em quatro tipos de acordo com as suas características morfológicas e comportamentais relacionadas as extremidades nervosas que serão vistas a seguir (Wyke, 1979; Alter, 1999).
Receptores articulares do Tipo I
Os mecanorreceptores do Tipo I consistem de corpúsculos globulares finamente encapsulados e localizam-se na camada externa das cápsulas fibrosas. Em cada corpúsculo globular é provido de uma fibra mielinizada do grupo II. São mais encontrados nas articulações proximais (quadril) do que articulações mais distais (tornozelo).
São caracterizados como receptores de limiar baixo, de adaptação lenta. Sendo assim, respondem a estresses mecânicos muito pequenos. Mesmo quando a articulação está imóvel, uma proporção desses receptores está sempre ativa (Wyke, 1979; Alter, 1999).
Receptores articulares do Tipo II
O receptor do tipo II consistem de corpúsculos maiores espessamente encapsulados e cônicos e localizam-se na cápsula articular fibrosa em camadas subsinoviais profundas. Também são mais localizados em maior densidade nas articulações mais distais do que nas articulações mais proximais.
Assim como os receptores do tipo I, os receptores do tipo II têm um limiar baixo, porém se adaptam mais rapidamente e não aquecem em repouso, sendo inativos nas articulações imóveis. Também são conhecidos como mecanorreceptores de aceleração ou dinâmicos e sua principal função é medir mudanças rápidas no movimento, como a aceleração e desaceleração (Wyke, 1979; Alter, 1999).
Receptores articulares do Tipo III
O receptor articular do tipo III são corpúsculos finamente encapsulados, confinados aos ligamentos intrínsecos (dentro da cápsula articular) e extrínsecos (fora da cápsula articular) de muitas articulações. São os maiores dos corpúsculos articulares e são como os órgãos tendinosos de Golgi, comportando-se como mecanorreceptores de alto limiar que se adaptam lentamente. São completamente inativos nas articulações imóveis e respondem somente quando altas tensões são geradas nos ligamentos de cada articulação (Wyke, 1979; Alter, 1999).
Receptores articulares do Tipo IV
As extremidades nervosas do tipo IV são desencapsuladas, portanto são subdivididas em dois tipos. O receptor do tipo IVa são representadas pelos plexos em forma de treliça, que são encontrados em grandes coxins articulares e por toda a espessura da cápsula articular, porém, com ausência de tecido sinovial, menisco intra-articular e cartilagem articular. Já o do tipo IVb são extremidades nervosas livres, são esparsos e largamente confinados aos ligamentos intrínsecos e extrínsecos (ALTER, 1999).
Ambos os tipos IVa e IVb constituem o sistema receptor de dor dos tecidos articulares, sendo chamados de nociceptores. Quando ocorre acentuada deformação mecânica ou irritação química, como por exemplo, ácido lático, íons potássio e histaminas, podendo aparecer em situações de isquemia (falta de sangue) e hipóxia (falta de oxigênio) (Wyke, 1979; Alter, 1999).
Reflexo miotático ou de alongamento
Quando um músculo é alongado, o mecanismo reflexo de alongamento é iniciado, estiram-se as fibras musculares extrafusais e os fusos musculares e os potenciais de ação são estimulados, o que acarretará em um impulso transmitido pelo axônio resultando em uma contração reflexa que pode ser dividida em dois componentes: fásico e tônico (ALTER, 1999).
A resposta fásica seria uma grande ocorrência de potenciais de ação resultando em uma elevação rápida da tensão muscular, proporcional à velocidade de alongamento, como por exemplo, a contração do joelho ou reflexo patelar. Já a resposta tônica seria uma fase posterior de estimulação lenta persistente por toda a duração do alongamento, sendo proporcional a quantidade de alongamento, como por exemplo, na reação postural ao alongamento na contração do músculo gastrocnêmio (panturrilha) para corrigir um movimento para frente excessivo do centro de gravidade de um indivíduo enquanto se levanta (ALTER, 1999).
Inibição autógena
Inibição autógena é o relaxamento neurológico de um músculo quando estimulado. Poderá ocorrer quando for ativado o OTG, gerando impulsos que irão proteger os músculos inibindo os neurônios motores α, relaxando os músculos (Gordon & Ghez, 1991; Andrews et al., 2005).
Inibição recíproca
A inibição recíproca inibe o músculo antagonista quando o músculo agonista move um membro na amplitude do movimento (BANDY & SANDERS, 2003), ou seja, os músculos geralmente atuam em pares, sendo que quando os agonistas contraem, os músculos opostos, os antagonistas, relaxem (ALTER, 1999; ANDREWS et al., 2005). Durante a flexão ativa do quadril, por exemplo, a inibição recíproca relaxa os músculos isquiotibiais. Esse relaxamento possibilita que os flexores do quadril possam realizar o movimento sem serem influenciados pelos músculos isquiotibiais em contração (GORDON & GHEZ, 1991). Em resumo, quando os motoneurônios de um músculo recebem impulsos excitatórios que levam à contração muscular, os motoneurônios do músculo oposto recebem sinais neurais, o que torna menos provável que eles sejam estimulados e produzam contração muscular (inibição).
Coativação ou co-contração
A coativação ou co-contração é a contração de dois músculos opostos com um alto nível de atividade nos músculos agonistas, simultaneamente com um baixo nível de atividade no músculo antagonista da mesma articulação (Solomonow & D’Ambrosia, 1991; Alter, 1999). Preserva a estabilidade articular, tendo o efeito mecânico de fazer uma articulação mais rígida, tornando o movimento mais difícil (ENOKA, 1988).
Fatores limitantes da flexibilidade
Tecido Conjuntivo
O tecido conjuntivo contém uma enorme variedade de células especializadas, que realizam funções de defesa, proteção, armazenamento, transporte, ligação, conexão e suporte geral e reparo (ALTER, 1999).
Colágeno
O colágeno é uma proteína que contém três cadeias de aminoácidos enrolados em uma hélice tripla em abundância em nosso corpo, tendo como as duas principais propriedades físicas das fibras colágenas de grande força de tração e relativa inextensibilidade. As fibras colágenas são capazes de um leve grau de extensibilidade, sendo os principais constituintes dos ligamentos e tendões que são submetidos a uma força de tração (ALTER, 1999).
O colágeno de um tendão é arranjado em feixes ondulados chamados fascículos, que são compostos de feixes de fibrilas, que são compostas de feixes de subfibrilas de colágeno que contém as microfibrilas ou filamentos de feixes de colágeno (KASTELIC et al., 1978).
A microfibrila colágena é composta de moléculas de colágeno espaçadas regularmente e sobrepostas, sendo análogas aos sarcômeros das células musculares. Elas se estendem em alinhamento paralelo com uma sobreposição alternada de quase um quarto de seu comprimento (ALTER, 1999). Um aspecto importante que ajuda na força de tração para as estruturas colagenosas é a presença de elos cruzados intramoleculares entre as cadeias α1 e α2 da molécula de colágeno e de elos cruzados intermoleculares entre as subfibrilas de colágeno, filamentos e outras fibras. Esses elos cruzados agem para unir as moléculas em uma unidade forte em forma de corda. Sendo assim, quanto menor for o comprimento entre um elo cruzado e o próximo ou quanto maior o número de elos cruzados em uma determinada distância, maior será a elasticidade (ALEXANDER, 1988).
O colágeno está sendo sempre produzido e decomposto, ou seja, se a produção exceder a decomposição, mais elos cruzados serão estabelecidos e a estrutura torna-se mais resistente ao alongamento. O exercício físico ou a mobilização podem reduzir o número de elos cruzados, aumentando a taxa de renovação do colágeno (Shephard, 1982; Alter, 1999). O colágeno envelhece, pois mudanças físicas e bioquímicas acontecem, reduzindo a extensibilidade e aumentando a rigidez.
Tecido elástico
O tecido elástico desempenha um papel importante na determinação da amplitude possível de extensibilidade das células musculares, desenvolvendo uma variedade de funções, como disseminar estresses, aumentar a coordenação dos movimentos rítmicos, armazenar energia, mantendo a tonicidade durante o relaxamento dos elementos musculares, desenvolvendo defesa contra forças excessivas (Jenkins & Little, 1974).
A elastina é uma estrutura complexa com uma propriedade mecânica de elasticidade devido a sua composição bioquímica e ao arranjo físico de suas moléculas. Com o envelhecimento, elas perdem a sua elasticidade, aumentam a sua rigidez e sofrem várias outras alterações, como fragmentação, desgaste, calcificação e outras mineralizações e um grande número de ligações cruzadas (Gosline, 1976; Alter, 1999).
Em relação ao limite fisiológico do alongamento da elastina, somente quando as fibras elásticas são alongadas até aproximadamente 150% de seu comprimento original, alcançam o seu ponto de quebra (Bloom & Fawcett, 1986; Alter, 1999).
Tendões
Os músculos são conectados aos ossos por cordões rígidos chamados tendões, tendo como principal função a transmissão da tensão do músculo para os ossos, produzindo o movimento. Tem como principais componentes, os fascículos colagenosos paralelos firmemente comprimidos que variam em comprimento e espessura. O tendão é adaptado para resistir ao movimento em uma direção. Portanto, quanto maior a proporção de colágeno para as fibras elásticas, maior será o número de fibras que são orientadas na direção do estresse e, quanto maior a área transversal ou a largura do tendão, mais forte será o mesmo (ALTER, 1999). Um estresse de aproximadamente 4% é importante e corresponde ao limite de elasticidade, nesse caso a ondulação da superfície do tendão desaparece e, se o alongamento continuar, pode resultar em lesão (CRISP, 1972).
Ligamentos
Os ligamentos ligam osso com osso, tendo como principal função a sustentação de uma articulação. Sendo muito semelhantes aos tendões, porém os tendões são compostos de feixes de fibras colagenosas situadas em paralelo ou entrelaçadas umas com as outras e os ligamentos são encontrados como cordas, bandas ou folhas, sendo maleáveis e flexíveis com o intuito de dar liberdade ao movimento, porém fortes, inextensíveis e rígidos para não ceder facilmente às forças aplicadas (ALTER, 1999).
Fáscia
A fáscia é todas as estruturas conectivas fibrosas, que varia em espessura e densidade de acordo com as exigências funcionais e encontra-se, geralmente, na forma de folhas membranosas. Pode ser dividida em três tipos: a fácia superficial (pele), é composta de duas camadas, a camada externa é denominada de panículo adiposo e a camada interna é uma membrana fina que geralmente não tem gordura; a fáscia profunda está diretamente abaixo da fáscia superficial, normalmente mais rígida, mais firme e mais compacta que a fáscia superficial além de proteger e dividir o corpo separando os músculos e os órgãos viscerais internos; a fáscia suberosa é mais profunda em volta das cavidades do corpo formando a camada fibrosa das membranas serosas que cobrem e sustentam a víscera (pleura que reveste os pulmões) (ALTER, 1999).
A fáscia tem como função, fornecer uma estrutura que liga o músculo e assegura o alinhamento adequado das fibras musculares, vasos sangüíneos e nervos, além de permitir que as forças sejam transmitidas por todo o tecido de forma segura e eficaz e também fornece as superfícies lubrificadas necessárias entre as fibras musculares e os feixes de fibra muscular que permitem que o músculo mude sua forma (ALTER, 1999).
Conclusão
A flexibilidade pode ser influenciada pelo sexo, idade, nível de treinamento, raça, histórico de lesão, reflexos espinhais, a temperatura, a hora do dia e a composição do músculo. Dois mecanorreceptores importantes são os fusos musculares e os órgãos tendinosos de Golgi, e os receptores articulares sentem forças mecânicas nas articulações como a pressão de alongamento e distensão. Ainda, o tecido conjuntivo é influenciado pelo envelhecimento, imobilização, distúrbios metabólicos e deficiências nutricionais, e por ser um dos componentes mais influentes na limitação da amplitude do movimento. Enfim, espera-se que esta revisão estimule o desenvolvimento de novas pesquisas que possam contribuir para o conhecimento dos profissionais da área da saúde nos aspectos fisiológicos da flexibilidade e os seus fatores limitantes.
Referências
-
ALEXANDER, R.M. Elastic mechanisms in animal movement. Cambridge: Cambridge University Press, 1988.
-
ALTER, M.J. Ciência da Flexibilidade. 2ed., Porto Alegre, Artmed, 1999.
-
ANDREWS, J.R.; HARRELSON, G.L.; WILK, K.E. Reabilitação Física do Atleta. 3ed., Rio de Janeiro, Elsevier, 2005.
-
BANDY, W.D.; SANDERS, B. Exercício Terapêutico: Técnicas para Intervenção. Rio de Janeiro, Guanabara Koogan, 2003.
-
BLOOM, W.; FAWCETT, D.W. A textbook of histology. Philadelphia: W.B.Saunders, 1986.
-
CRISP, J. Properties of tendon and skin. In: YUNG, Y.C.; PERRONE, N.; ANLIKER, M. (Eds.), Biomechanics: Its foundation and objectives, 141-180, 1972.
-
ENOKA, R.M. Neuromechanical basis of kinesiology. Champaign, IL: Human Kinetics, 1988.
-
FRONTERA, W.R.; DAWSON, D.M.; SLOVIK, D.M. Exercício físico e reabilitação. Porto Alegre, Artmed, 2001.
-
GORDON, J.; GHEZ, C. Muscle receptors and spinal reflexes: the stretch reflex. In: ER KANDEL, JH SCHWARTZ. Principles of neural science. 3ed. New York: Elsevier;564-580, 1991.
-
GOSLINE, J.M. The physical properties of elastic tissue. In: HULL, D.A.; JACKSON, D.S. International Review of Connective Tissue Research, 7:211-257, 1976.
-
GUYTON, A.C.; HALL, J.E. Textbook of Medical Physiology. 9ed., EUA, Saunders Company, 1996.
-
KASTELIC, J.; GALESKI, A.; BAER, E. The multicomposite structure of tendon. Connective Tissue Research, 6(1):11-23, 1978.
-
JENKINS, R.; LITTLE, R.W. A constitutive equation for parallel-fibered elastic tissue. Journal of Biomechanics, 7(5):397-402, 1974.
-
MONTEIRO, G.A. Treinamento da Flexibilidade: Sua aplicabilidade para Saúde. Londrina, Midiograf, 2006.
-
SHEPHARD, R.J. Physiology and biochemistry of exercise. New York: Praeger, 1982.
-
SOLOMONOW, M.; D’AMBROSIA, R. Neural reflex arcs and muscle controlo f knee stability and motion. In: SCOTT, W.N. (Ed.), Ligament and extensor mechanism injuries of the knee, (389-400), 1991.
-
WILMORE, J.H.; COSTILL, D.L. Fisiologia do Esporte e do Exercício. Barueri, Manole, 2001.
-
WYKE, B. Neurology of the cervical spinal joints. Physioterapy, 65(3):72-76, 1979.
Outros artigos em
Portugués
| |

Búsqueda personalizada
|
|---|---|
|
EFDeportes.com, Revista
Digital · Año 15 · N° 148 | Buenos Aires,
Septiembre de 2010 |
|