
|
|||
|---|---|---|---|
|
|
O metabolismo da amônia durante o exercício
Ammonia metabolism during exercise |
|
|
|
Laboratório de Pesquisa Morfo - Funcional. Centro de Educação Física Fisioterapia e de Desportos, CEFID. Universidade do Estado de Santa Catarina - UDESC. Florianópolis/SC. (Brasil) |
Vitor Pereira Costa Tony Charles Fernandes Monique da Silva Gevaerd Loch costavp@nextwave.com.br |
|
|
|
|
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 12 - N° 114 - Noviembre de 2007 |
|
|
1 / 1
Introdução
A amônia é um produto do metabolismo celular, sendo encontrada na forma não dissociada NH3 e íons NH+4 em solução fisiológica (GRAHAM et al., 1997). A amônia, aqui tratada de NH3, é produzida em diversas células no fígado, músculos, intestino, tecido nervoso, rins e eritrócitos (MUTCH & BANISTER, 1983). Durante o exercício, o músculo é considerado o principal produtor de NH3 através do Ciclo da Purina Nucleotídeo (CPN) e pelo consumo de aminoácidos de cadeia ramificada (BCAA), (GRAHAM et al., 1997). Os mecanismos de produção, estão intimamente relacionados com as reações bioquímicas do metabolismo energético contudo ainda não completamente esclarecidos.
Existem diferentes tecidos que podem sofrer alterações agudas com a alta toxidade de NH3. Altos níveis circulantes de NH3 pode ser tóxica tanto para o músculo quanto para o sistema nervoso central, resultando em funções neurais alteradas, confusão mental, desorientação, alucinação, ataque epilético, convulsões, coma e hiperpnéia (MUTCH & BANISTER, 1983; BANISTER & CAMERON, 1990). Entretanto, as modificações ocorridas durante o exercício parecem ser mais discretas e menos prejudiciais ao organismo humano, exceto em casos extremos (BANISTER & CAMERON, 1990).
Elevadas concentrações de NH3, também podem levar a alterações metabólicas que estimulam a enzima fosfofrutoquinase e inibem a piruvato e a isocitrato desidrogenase (MUTCH & BANISTER, 1983). A fosfofrutoquinase é uma enzima chave que regula a velocidade da glicólise anaeróbia onde o produto final desta via pode ser lactato ou piruvato. Sendo assim, altos níveis de NH3 podem estar associados com altos níveis de produção de lactato (YUAN et al., 2002). Além disso, alguns estudos sugerem que a NH3 está relacionada com a fadiga central e periférica (BANISTER & CAMERON, 1990).
Devido à abrangência de trabalhos sobre os mecanismos da síntese e utilização da NH3 e suas possíveis relações com aspectos clínicos, o presente estudo tem como objetivo descrever sobre o metabolismo da NH3 e suas implicações durante o exercício. Sendo assim, esta breve revisão organiza as diferentes etapas no processo de produção e as inferências sugeridas com o exercício.
Produção de NH3 durante o exercícioDurante o exercício ocorre um aumento das concentrações musculares e sanguíneas de NH3. A intensidade e a duração do esforço parecem estar relacionadas diretamente com o aumento da produção de NH3 no músculo esquelético (GRAHAM et al., 1997).
Em esforço intenso, o ritmo na utilização de ATP no músculo esquelético, é mais rápido do que sua regeneração, o que pode levar no acúmulo de ADP e AMP. Para evitar grande acúmulo dentro da célula, o AMP é desaminado em IMP e NH3, diretamente pela atividade da enzima AMP desaminase.
A NH3 também pode ser formada através do Ciclo da Purina Nucleotídeo (CPN) (LOWESTEIN, 1990). O CPN ocorre no sarcoplasma da fibra muscular e consiste em reações catalisadas por três enzimas específicas, AMP desaminase, adenilsuccinato sintetase e adenilsuccinato liase (Figura 1). A primeira reação (a), consiste na hidrólise do AMP com a atuação da enzima chave AMP desaminase, resultando na formação de uma molécula de IMP e NH3. No segundo passo (b), o IMP intramuscular pode ser reduzido pela combinação com aspartato e formar adenilsuccinato. Esta reação é catalisada pela adenilsuccinato sintetase com consumo de energia (GTP). Em seguida (c), o produto é convertido em AMP e fumarato, pela adenilsuccinato liase (LOWESTEIN, 1990).
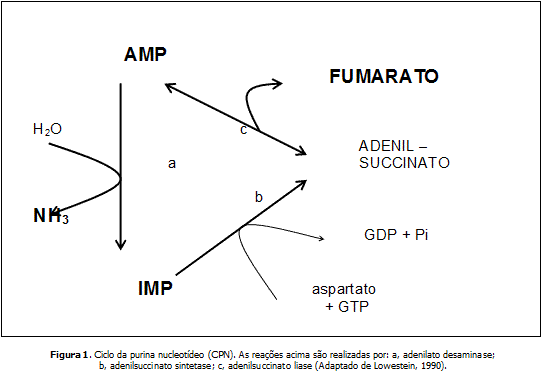
As funções relacionadas com a ativação do CPN participam na manutenção das concentrações celulares de AMP, ADP e ATP, em particular, manter elevada a razão ATP/ADP. Estes compostos atuam como substratos, ativadores e inibidores de muitas reações, de forma geral, participam no controle metabólico. O CPN auxilia também na regulação do metabolismo energético pois elevadas concentrações de AMP parecem ser um importante modulador alostérico da fosforilase ativa (importante enzima que cataboliza o glicogênio muscular) e fosfofrutoquinase, com o conseqüentemente aumento da glicólise anaeróbia. Em adição, durante o exercício severo os níveis de IMP se elevam e alteram a atividade da enzima fosforilase b. Assim, o ciclo desempenha funções na regulação celular, consumo de aninoácidos (AA), eliminação da NH3 e manutenção de intermediários do ciclo do ácido cítrico.
Além dos eventos que ocorrem no sarcoplasma, a formação de NH3 pode ser proveniente do catabolismo de aminoácidos de cadeia ramificada (BCAA) nas mitocôndrias em exercícios de longa duração (GRAHAM et al., 1997). De forma simplificada, os BCAA podem ser transaminados a seus respectivos cetoácidos, com o alfacetoglutarato recebendo o grupo amino, para formar o glutamato. Os cetoácidos podem ser transportados ao fígado para futura oxidação ou serem catabolisados no músculo esquelético. Em ambos os casos, a enzima responsável pela reação é a cetoácido desidrogenase que está presente em uma forma mais ativa e menos ativa. No músculo esquelético em repouso, especula-se que esta enzima se encontra em uma baixa atividade, assim, parte dos cetoácidos, podem ser transportados até o fígado para serem oxidados. No entanto, durante o exercício, os cetoácidos podem ser catabolisados no músculo (HARGREAVES, 1995). Ao final deste processo, podemos obter a liberação de NH3 livre pelo glutamato, tendo como origem no processo de.transaminação e desaminação oxidativa de AA.
De fato, o glutamato que, temporariamente armazena o grupo amino, pode ser consumido por diferentes vias durante o exercício prolongado. As vias metabólicas onde há grande consumo de glutamato durante o exercício ainda não foram completamente estabelecidos (GRAHAM et al., 1997). No entanto, é bem aceito que NH3 livre pode se combinar com o glutamato e formar glutamina e ser liberada do músculo. A glutamina é um dos principais transportadores de NH3 no sangue para a desaminação oxidativa hepática. O glutamato também pode ser transaminado com o piruvato para formar alanina e participar na gliconeogênese através do ciclo glicose-alanina liberando o grupo amino no fígado. O glutamato também pode ser transaminado com o oxaloacetato e formar aspartato. É importante lembrar que o aspartato, além de participar na síntese de uréia, também funciona como substrato do CPN. Em adição, o glutamato pode sofrer desaminação oxidativa pela glutamato desidrogenase e produzir NH3 livre (HARGREAVES, 1995).
Assim, fica evidente a possibilidade que todos esses processos na formação de NH3 podem ocorrer durante o exercício. As investigações que objetivam reconhecer a origem da NH3 na musculatura esquelética precisam considerar os caminhos supracitados. No entanto, não está claro se o consumo de glutamato intramuscular envolve processos de transaminação ou na formação de glutamina. Em adição, os mecanismos responsáveis pela desaminação oxidativa foram reconhecidos no tecido hepático, entretanto, no tecido muscular necessita de melhores explicações. Outra questão envolve a formação de alanina durante o exercício intenso, um dos substratos da reação é o piruvato que pode estar sendo consumido em outras vias.
Inferências sugeridas com o exercícioAs medidas das concentrações sanguíneas [NH3] têm se destacado na literatura científica no que concerne a aspectos de desempenho no esporte (GRAHAM et al., 1997; YUAN et al., 2004). As [NH3] e o limiar da amônia (LA) foram investigados na tentativa de consolidarem como novas referências no treinamento esportivo (HAGELOCH et al., 1990; GRAHAM et al., 1997; YUAN et al., 2004). Sendo assim, foram selecionados alguns estudos publicados até o presente momento, sobre as especulações relacionadas com o metabolismo da NH3 e sua influência durante o exercício.
Exercícios anaeróbiosA produção de NH3 no músculo esquelético depende dos tipos de fibras musculares com apropriadas atividades enzimáticas envolvidas. Meyer e Terjung, (1979) verificaram que as fibras de contração rápida apresentam grande atividade da AMP desaminase, enquanto que fibras de contração lenta possuem menor atividade desta enzima. Sahlin et al., (1989) investigaram que após o exercício dinâmico, o IMP foi formado em ambos os tipos de fibras, mas as concentrações foram mais altas nas fibras de contração rápida que as de contração lenta. Em adição, as concentrações de IMP provenientes de análises musculares, são consideradas como um fidedigno marcador local de produção de NH3, visto que o IMP não pode se difundir facilmente pelo sarcolema (MEYER et al., 1980).
As diferenças do metabolismo da NH3 durante o exercício estão relacionadas em parte, com a intensidade, duração do esforço e intimamente com os diferentes tipos de fibras musculares. Durante o exercício intenso, a razão de ATP/ADP se torna reduzida com acúmulo e posterior desaminação de AMP. Em adição, fibras de contração rápida são recrutadas resultando em maior atividade do CPN, conseqüentemente maior produção de NH3 (MEYER e TERJUNG, 1979). No entanto, em atividades de baixa intensidade ocorre grande disponibilidade de ATP, com menor formação de ADP e AMP. Assim, unidades motoras de contração lenta são predominantemente recrutadas, resultando em uma pequena produção de NH3 (MEYER e TERJUNG, 1979).
Elevadas concentrações de NH3 podem estar associados com altos níveis de produção de lactato. (YUAN et al., 2002). Na tentativa de descrever um novo parâmetro metabólico no desporto, Schlicht et al., (1990) analisaram diferentes informações entre a produção de NH3 e lactato. Os autores verificaram que após a realização de quatro corridas em intensidade correspondente a 87,5% do pico de velocidade durante 300 ms de percurso, as [NH3] aumentam significativamente se comparada aos valores de repouso. Em contraste, as [La] começaram a aumentar, na primeira sessão de corridas a 82% do pico de velocidade. De fato, NH3 e lactato, podem trazer diferentes informações sobre o metabolismo muscular e são ativados entre 80 e 90% do pico de velocidade, respectivamente.
De acordo com esta linha, Hageloch et al. (1990) tentaram verificar se as [NH3] identificadas em teste específico de campo, podem servir como indicadores na seleção de futuros talentos nas modalidades de potência anaeróbia. Os autores avaliaram jovens velocistas e corredores de meio fundo, durante três corridas realizadas na máxima velocidade possível, a 25, 75 e 1000 ms. Após o teste de 75 ms, os velocistas apresentaram os maiores valores de [NH3], enquanto que após 1000 ms, os corredores de meio fundo apresentaram maior pico de [NH3]. Em contraste, as [La] não apresentaram diferenças significativas entre os grupos, apesar de demonstrar uma ligeira tendência em ser mais elevada nos corredores de meio fundo. A partir dos resultados, os autores sugerem que a razão de [NH3] após o teste de 75 /1000 ms, pode ser definida como um índice, em que valores mais altos seriam típicos de atletas velocistas. Apesar da sugestão, existem críticas, por exemplo, em situações onde o índice poderia ser aplicado em mulheres ou em adultos, e sobre a influência de outros métodos de treinamento.
De fato, não se pode negar que existe uma relação entre produção de NH3 e lactato em exercícios anaeróbios. É bem sugerido que a relação entre o aumento de lactato intramuscular e o ritmo de desaminação do AMP, ocorre pela ativação direta da AMP desaminase e pelos íons H (SAHLIN e BROBERG, 1990). Em adição, o aumento dos níveis de ADP e AMP pode estimular a PFK em manter a velocidade da glicólise mesmo com a queda do Ph. Assim o AMP pode ser desaminado e resultar no aumento da formação de IMP e NH3 (SAHLIN e BROBERG, 1990). Neste sentido, fica evidente a interação entre a produção de NH3 e lactato, no entanto, a dissociação entre essas variáveis identificadas nos diferentes modelos apresentados por Schlicht et al. (1990) e Hageloch et al. (1990) não podem ser respondidas facilmente.
Exercícios aeróbiosOs limiares ventilatórios (LV) e de lactato (LL) têm sido utilizados como referência de capacidade aeróbia no treinamento desportivo com o intuito de otimizar o desempenho dos atletas (LUCIA et al., 2002; COSTA et al., 2005). Na realidade, esses limiares representam momentos de transição metabólica, impostas ao sistema energético a partir da demanda de trabalho exigida. Nos últimos anos, as [NH3], por também serem um produto metabólico, tem sido referenciada em algumas situações, por apresentar o mesmo fenômeno de transição como ocorre com os limiares (BUONO et al., 1984). No entanto, a concomitante similaridade entre o LA e LL, parece acontecer em momentos distintos (BANISTER et al., 1983).
Banister et al. (1983) utilizaram protocolo de rampa no ciclo-ergômetro e verificaram que as [NH3] aumentam a partir de 40 - 45 % do VO2máx. Em adição, as [NH3] atingem o pico e diminuem rapidamente. Em contraste, as [La] aumentam sistematicamente, apenas a partir de 80 % do VO2máx, sendo que diminuem lentamente. De fato, observa-se a dissociação entre o tempo de aparecimento e remoção das [NH3] e [La]. Dentre as possíveis explicações, considera-se o ritmo distinto de efluxo de cada um dos metabólitos provenientes do tecido exercitado (BANISTER et al., 1983).
Recentemente, Yuan et al. (2002) compararam as intensidades referentes ao LA e LL, em cinco avaliações diferentes. Os resultados indicam que apenas o protocolo contínuo com estágios de 4 min, apresenta a intensidade de esforço distinta entre os limiares (p<0,01). Especula-se que estágios com longa duração, podem afastar os limiares investigados, pois são obtidas poucas coletas. No entanto, os autores apresentaram que em protocolo de estágios de 8 min, não houve diferença significante entre as intensidades nos LL e LA.
De fato, a semelhança entre os limiares encontrados nos diferentes protocolos contínuos e intermitentes apresentados por Yuan et al. (2002), corroboram com os achados de Buono et al. (1984) que verificaram associação entre a produção de NH3 e o metabolismo glicolítico durante o exercício. No entanto, é importante ressaltar que a identificação do LA, talvez seja confirmada, apenas em algumas situações específicas, que em grande parte, ainda estão limitadas a protocolos de cargas progressivas, distante das situações onde se verifica o máximo estado de equilíbrio das variáveis de capacidade aeróbia.
A problemática em relação a identificação do LA e LL não se limita apenas a protocolo-dependência. Os diferentes métodos de identificação de limiares de transições metabólicas, muitas vezes, causam dúvidas e são difíceis de serem interpretados. Em grande parte, esses métodos, objetivam se aproximar do máximo estado de equilíbrio do lactato sangüíneo. Assim, Yuan et al. (2002; 2004) e Langfort et al. (2004) identificam os limiares a partir da intersecção de dois segmentos, sendo que este protocolo não representa necessariamente, o máximo estado de equilíbrio das variáveis estudadas.
Em adição, Langfort et al (2004) apresentaram que as modificações dietéticas também parecem contribuir para a dissociação entre a identificação dos limiares metabólicos, sendo que uma dieta reduzida em carboidratos diminui a velocidade da glicólise e conseqüentemente a produção de lactato, por outro lado, aumenta NH3. Neste sentido, Snow et al. (2000) indicam que a ingestão de carboidratos durante o exercício a 65 % do VO2máx , atenua o acúmulo plasmático e muscular de NH3 durante os últimos estágios do esforço.
Em protocolo de cargas constantes, Ogino et al. (2001) verificaram as respostas de [NH3] e [La] no ciclo-ergômetro, em diferentes intensidades correspondentes a 80%, 90%, 100%, 110% e 120% do LV, durante 15 minutos. Os autores apresentaram que as [NH3], aumentam a partir do sexto minuto de esforço e durante todas as cinco intensidades de exercício. Em contraste, as [La] não aumentam continuamente em intensidades abaixo do LV. Corroborando, MacLean et al. (1991) verificaram as respostas plasmáticas e musculares de NH3 e AA durante o exercício prolongado. Os resultados também indicam discordância entre a elevada produção de NH3 e estabilidade na produção de lactato.
Assim, uma possível explicação para a elevada produção de NH3 nas cargas inferiores ao LV, baseia-se em que exercícios de baixa intensidade, os AA podem promover uma fonte alternativa na produção de NH3 (VAN HALL et al., 1995; GRAHAM et al.,1997). No entanto, a participação de AA como fonte de substrato energético se torna mais evidente em exercícios de longa duração e de força (BROOKS, 1987; TIPTON et al., 1999).
Um adicional argumento para a limitada atividade da enzima AMP desaminase e do CPN durante o exercício sub-máximo, é proveniente do recrutamento de fibras musculares (MacLEAN et al., 1991). A atividade da AMP desaminase e a produção de IMP, foram significantemente elevadas nas fibras musculares de contração rápida (MEYER e TERJUNG, 1979). Durante o exercício prolongado de intensidade a 75% do VO2máx , ocorre a predominância no recrutamento de fibras musculares de contração lenta, com algumas fibras de contração rápida sendo recrutadas próximas a exaustão (GOLLNICK et al., 1974). Como resultado, a atividade da AMP desaminase durante grande parte do exercício parece ser reduzida.
Contudo, não se pode dizer que no exercício prolongado, não ocorre a participação do CPN, de fato, os estudos apresentam que a produção de NH3 continua a se elevar na medida em o esforço se prolonga (MacLEAN et al., 1991; OGINO et al., 2001). Neste sentido, além da participação do CPN, os AA de uma forma geral, parecem participar no metabolismo da NH3 durante o exercício (MacLEAN et al., 1991; VAN HALL et al., 1995; GRAHAM et al., 1997).
Considerações finaisAssim, percebe-se que os processos envolvidos na produção de NH3 durante o exercício são complexos e até o presente momento ainda não foram completamente esclarecidos. Acredita-se, que a intensidade e a duração do esforço, estejam relacionados com o aumento das concentrações sanguíneas e intramusculares de NH3. No entanto, talvez a quantidade de massa muscular envolvida, os diferentes tipos de exercício e o nível de aptidão física, podem promover diferentes respostas nos mecanismos de formação. Diversos autores sugerem que as [NH3] podem servir como referências do treinamento de modalidades aeróbias e anaeróbias. Entretanto, são necessários novos estudos que utilize esta variável para avaliação, prescrição e controle, bem como os efeitos do treinamento.
AgradecimentoAgradecemos a contribuição do Dr. Fernando Roberto de Oliveira, que nos auxiliou com revisão crítica desta obra.
Referências bibliográficas
BANISTER, E. W.; ALLEN, M. E.; MEKJAVIC, I. B.; SINGH, A. K.; LEGGE, B.; MUTCH, B. J. C. The time course of ammonia and lactate accumulation in blood during bicycle exercise. European Journal of Applied Physiology. 51: 195 - 202, 1983.
BANISTER, E. W.; CAMERON, B. J. C. Exercise-induced hyperammonemia: peripheral and central effects. International Journal of Sports Medicine, Stuttgart, 11: S129-S142, 1990.
BROOKS, G. A. Amino acid and protein metabolism during exercise and recovery. Medicine and Science in Sports and Exercise, Madison, 19: S150 - S156, 1987.
BUONO, M. J.; CLANCY, T. R.; COOK, J. R. Blood lactate and ammonium ion accumulation during graded exercise in humans. Journal of Applied Physiology, Bethesda, 57: 135-139, 1984.
COSTA, V. P; FERNANDES, T. C.; ADAMI, F.; CARMINATTI, L. J.; LIMA-SILVA, A. E.; DE-OLIVEIRA, F. R. Ponto de inflexão como estimativa dos limiares de lactato em mountain bikers. Anais do I Encontro Mineiro de Fisiologia do Exercício. Juiz de Fora, UFJF, 2005.
GOLLNICK, P.; PIEHL, K.; SALTIN, B. Seletive glycogen depletion pattern in human muscle fibers after exercise of varying intensity and varying pedal rates. Journal of Physiology, London, 241: 45-58, 1974.
GRAHAM, T. E.; TURCOTTE, L. P.; KIENS, B.; RICHTER, E. A. Effect of endurance training on ammonia and amino acid metabolism in humans. Medicine and Science in Sports and Exercise, Madison, 29(5): 646 - 53, 1997.
HAGELOCK, W.; SCHNEIDER, S.; WEICKER, H. Blood ammonia determination ina a specific field test as a method supporting talent selection in runners. International Journal of Sports Medicine Stuttgart, 11: S37-S46, 1990.
HARGREAVES, M. Exercise Metabolism. Champaign IL: Human Kinetics, 1995;
LANGFORT, J.; CZARNOWSKI, D.; ZENDZIAN-PIOTROWSKA, M.; ZARZECZNY, R.; GORSKI, J. Short-term low-carbohydrate diet dissociates lactate and ammonia thresholds in men. Journal of Strength and Conditioning and Research, Champaign,18(2): 260-5, 2004.
LOWENSTEIN, J. M. The purine nucleotide cycle revised. International Journal of Sports Medicine, Stuttgart, 11 S37 - S46, 1990.
LUCIA, A.; JOYOS, H.; et al. Kinetics of VO2 in professional cyclists. Medicine and Science in Sports and Exercise, Madison, 34 (2): 320-325, 2002.
MEYER, R. A.; TERJUNG, R. L. Differences in ammonia and adenylate metabolism in contracting fast and slow muscle. American Journal of Physiology. 237: C111-C118, 1979.
MEYER, R. A.; DUDLEY, G. A.; TERJUNG, R. L. Ammonia and IMP in different skeletal muscle fibers after exercise in rats. Journal of Applied Physiology, Bethesda, 49: 1037-1041, 1980.
MacLEAN, D. A; SPRIET, L. L; HULTMAN, E; GRAHAN, T. E. Plasma and muscle amino acid and ammonia responses during prolonged exercise in humans. Journal of Applied Physiology, Bethesda, 70 (5): 2095 - 2103, 1991.
MUTCH, B. J. C; BANISTER, E. W. Ammonia metabolism in exercise and fatigue: a review. Medicine and Science in Sports and Exercise, Madison, 15 (1): 41 - 50, 1983.
OGINO, K.; KINUGAWA, T.; OSAKI, S.; et al. Ammonia response to constant exercise: differences to the lactate response. Clinical and Experimental Pharmacology and Physiology. 27(8): 612-617, 2000.
SAHLIN, K; BROBERG, S; REN, J. M. Formation of inosine monophosphate (IMP) in human skeletal muscle during incremental dynamic exercise. Acta Physiologyca Scandinavica. 136: 193 - 198, 1989.
SAHLIN, K.; BROBERG, S. Adenine nucleotide depletion in human muscle during exercise: causality and significance of AMP deamination. International Journal of Sports Medicine, Stuttgart, 11: S62-S67, 1990.
SCHLICHT, W.; NARETZ, W.; WITT, D.; RIECKERT, H. Ammonia and lactate: differential information on monitoring training load in sprint events. International Journal of Sports Medicine, Stuttgart, 11(2): S85-S90, 1990.
SNOW, R. J.; CAREY, M. F.; STATHIS, C. G.; FEBBRAIO, M. A.; HARGREAVES, M. Effect of carbohydrate ingestion on ammonia metabolism during exercise in humans. Journal of Applied Physiology, Bethesda, 88: 1576-1580, 2000.
TIPTON, K. D.; FERRANDO, A. A.; PHILLIPS. S. M.; DOYLE, D. Jr.; WOLFE, R. R. Post exercise protein net synthesis in human muscle from orally administered amino acids. American Journal of Physiology Endocrinology and Metabolism. 276: E628-E634, 1999.
VAN HALL, G; SALTIN, B; VAN DER VUSSE, G. J; SÖDERLUND, K; WAGENMAKERS, A. J. M. Deamination of amino acids as a source for ammonia production in human skeletal muscle during prolonged exercise. Journal of Physiology, London, 489 (1): 151 - 261, 1995.
YUAN, Y; SO, R; WONG, S; CHAN, K. M. Ammonia threshold - comparison to lactate threshold, correlation to other physiological parameters and response to training. Scandinavian Journal of Medicine and Science in Sports. 12: 358 - 364, 2002.
YUAN, Y; CHAN, K. M. A longitudinal study on the ammonia threshold in junior cyclists. British Journal of Sports Medicine, London, 38(2): 115 - 119, 2004.
 |
|
|---|---|
|
revista
digital · Año 12
· N° 114 | Buenos Aires,
Noviembre 2007 |
|